Latest News
- Clues beginning to emerge on asymtomatic SARS-CoV-2 infection
- Back in November of 2020, during the first wave of the COVID-19 pandemic, I was teaching an in-person microbiology laboratory. One of my students had just been home to see his parents, and they all c…
- Read more
- Could there maybe be better uses of genetics and probiotics?
- Professor Meng Dong and his laboratory have created a probiotic that can metabolize alcohol quickly and maybe prevent some of the adverse effects of alcohol consumption. The scientists cloned a highl…
- Read more
- ChatGPT is not the end of essays in education
- The takeover of AI is upon us! AI can now take all our jobs, is the click-bait premise you hear from the news. While I cannot predict the future, I am dubious that AI will play such a dubious role in…
- Read more
- Fighting infections with infections
- Multi-drug-resistant bacterial infections are becoming more of an issue, with 1.2 million people dying of previously treatable bacterial infections. Scientists are frantically searching for new metho…
- Read more
- A tale of two colleges
- COVID-19 at the University of Wisconsin this fall has been pretty much a non-issue. While we are wearing masks, full in-person teaching is happening on campus. Bars, restaurants, and all other busine…
- Read more
( 33069 Reads)
|Learning Objectives
- Describe the structure of antibodies and the 5 types that are found in the body.
- Describe the mechanisms used by the cell to create millions of antibodies.
- Explain how antibodies defend against pathogens.
B lymphocytes or B cells are the class of immune cells that synthesize antibodies and are responsible for humoral or antibody-mediated immunity. Antibodies react with antigens and the immune system has a virtually unlimited capacity to create antibodies that react with millions of potential antigens.
Each B cell contains on its surface about 100,000 identical antibody molecules (IgD or IgM) The binding sites of these antibodies will all react specifically to the same antigenic structure, but each unique B cell will contain a different antibody, reactive with a different antigen. These membrane-bound antibodies serve as receptors for the antigen and contain two additional polypeptides (dimers of a protein called Ig-α/Ig-β) that span the cellular membrane and play a role in transmitting the binding of antigen to antibody to the cytoplasm of the cell as shown in Figure 16.2. Binding of an antigen by several different antibody receptors on the surface of the cell causes the receptors to cluster in a small area. This clumping of membrane bound antibody molecules then activates a signaling cascade that eventually results in the transcription of genes and thus the production of proteins important for B cell activation.
Figure 16.2. B cell activation. Binding of antibody activates tyrosine kinase and tyrosine phosphorylase in the cytoplasm that then phosphorylates and dephosphorylates tyrosine residues on the Ig-α/Ig-β polypeptide. These phosphorylation/dephosphorylation reactions activate B cells by at least three different pathways: (i) Activated tyrosine residues on the Ig-α/Ig-β polypeptide cause phospholipase C activity to increase. Phospholipase C then cleaves phospholipids into inositol triphosphate and diacyl glycerol. Diacyl glycerol activates protein kinase C eventually leading to the formation of nuclear factor NF-κB. (ii) Inositol triphosphate causes Ca2+ influx from the endoplasmic reticulum and the outside environment. Increased Ca2+ concentrations activate calmodulin that in turn phosphorylates Ets-1, a DNA binding protein. (iii) Activated tyrosine residues on the Ig-α/Ig-β polypeptide activate the p21ras protein, which leads to the activation of a serine/threonine kinase. This in turn phosphorylates cJUN, another DNA binding protein. NF-κB, Ets-1 and cJUN then travel to the nucleus and cause the transcription of specific genes important in B cell activation.
The activated B cell then goes through a process of rapid division (termed clonal expansion) where many more copies of the cell are made. Some of these cells differentiate into plasma cells that produce large amounts of antibody. The antibody molecules produced specifically react with the antigen that caused the initial activation of the B cell. A fraction of the cells from the clonal expansion remain as undifferentiated B memory cells that are capable of reacting with a second antigen challenge at a later time. If a future challenge does appear, this secondary response is much more rapid, normally inactivating the pathogen before a detectable infection appears.
The “B" of B cell comes from the Bursa of Fabricius, where B cells differentiate in birds. (Some of the early research on the immune system was done in chickens). In mammals, B cells mature in the bone marrow and then travel in the bloodstream eventually settling in immune tissues throughout the body with high concentrations in the lymph nodes and spleen.
Antibodies Come in Five Different Classes
We will now spend a bit more time looking at the structure and function of antibodies to give you a more in-depth understanding of their different types and roles in the immune response.
Antibodies have the interesting challenge of needing to respond to a wide array of antigens, yet still be recognizable to the immune system. They therefore, need to interact with two different types of macromolecules, antigens and parts of the immune system. If antibodies were too similar to each other in structure, they would not be able to recognize many antigens, but if they were too different, they would not have a consistent enough structure to react with various immune components. Antibodies solve this dilemma by having two regions, a highly variable region that recognizes the antigens and constant regions that interact with the immune system.
Plasma cells synthesize a number of different types of antibodies and these serve different functions for the cell. Antibodies are proteins made up of two light chains and two heavy chains, see Figure 15.1. The heavy chain determines the type of antibody class and is bound to the light chain by sulfhydryl linkages. Each chain has a constant region that is similar amongst all antibodies. The constant region interacts with the rest of the immune system to facilitate the immune response. Each chain also has a variable region containing a high degree of amino acid variability that is unique to each antibody and this is the part that binds to the antigen. The proteins wrap together and form a Y structure with the two prongs of the Y containing the variable (Fab) regions and the bottom stem of the Y containing the constant (Fc) region.
The different classes of antibodies are defined by the structure of the constant region of the heavy chain. Each heavy chain gives the class of antibody certain properties. Here we first introduce the properties of these antibody classes and then describe the various roles antibodies play in the inactivation of pathogenic microorganisms.
Classes Of Antibodies
While billions of different variable regions are made, the general structure of antibodies falls into just five classes and this is based upon the type of heavy chain present in the antibody as shown in Figure 16.3. Immunoglobulin G (IgG) is the most abundant circulating antibody, making up 80% of the total antibodies and 75% of that found in serum. It contains a single antibody protein complex, with two heavy chains and two light chains. IgG is the second type of antibody synthesized in response to an infection and is the only antibody that can pass through the wall of small blood vessels to access antigens present in the extracellular spaces. It is also the only antibody capable of crossing the placenta in humans, where it confers the mother's immunity onto the fetus and newborn. This immunity protects a baby for the first 6-12 months of its life and allows it time for its own immune system to mature. IgG is particularly effective at attacking extracellular viruses and protein toxins and is also capable of activating the classic pathway of the complement cascade. It helps to prevent the systemic spread of infection and facilitates recovery from many infections. Finally, IgG is the antibody that serves as an efficient handle for phagocytes, allowing phagocytes to bind to a pathogen through Fc receptors on IgG and rapidly phagocytize a pathogen.
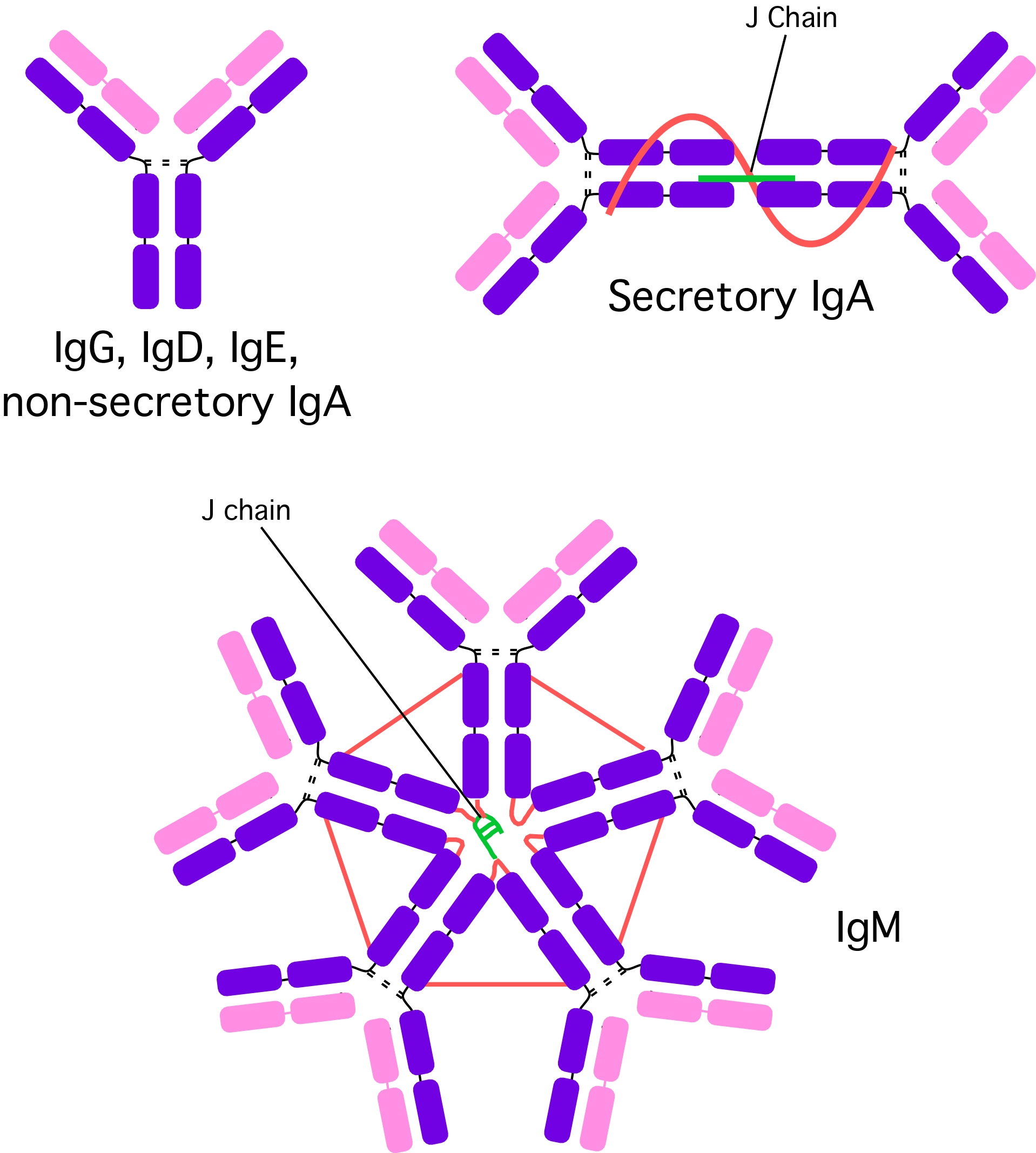
Figure 16.3. Different Antibody Types. There are five different antibodies classes that are formed in the cell. See the text for more details.
IgM is the largest antibody, with five Y structures being joined by their Fc regions in a circular configuration. A J chain (another polypeptide) links the five antibodies together. IgM is present in serum, making up about 10 % of antibodies in the blood. The presence of its ten antigen reactive sites helps agglutinate or clump antigens (see the explanation of this term in the next section), making it easier for the immune system to eliminate them. IgM is more efficient than IgG at activating the complement pathway. IgM is synthesized by plasma cells early in a primary infection and is very important in slowing or stopping the spread of a pathogen during the initial stages of an illness. IgM is also found on mature B cells in a monovalent form, where it serves as a receptor.
IgA is present in serum, mucus, saliva, tears, sweat and milk. Two subclasses with different heavy chains are made, IgA1 and IgA2. IgA1 is synthesized in the bone marrow and makes up most of the serum IgA. IgA2 is synthesized by B cells present in MALT. The antibodies are synthesized as dimers that are joined by a short J chain polypeptide. As the secreted IgA2 passes through the intestinal epithelium, a second secretory protein attaches. Dimerization and binding of the J and secretory proteins make IgA more resistant to proteases present in the environments that it protects. IgA in breast milk interferes with the colonization of the GI tract by harmful microorganisms in the first few months of life. The mother's IgA in the GI tract of newborns keeps these pathogens at low populations, preventing them from causing serious disease, but still allowing the stimulation of the infant's own immune system. The newborn thus develops its own immunity while being partially protected by the mother. IgA molecules do not activate the classical complement pathway, but may activate the alternative complement pathway.
IgE is a monomeric antibody that accounts for only 0.002 % of the total serum antibodies. Almost all IgE is bound to tissue cells, especially mast cells and eosinophils in various parts of the body. Contact of IgE with antigen leads to release of a set of signal molecules from the mast cells, which effectively recruits various agents of the immune response to fight the infection. IgE and MALT serve to detect penetrating pathogens and amplify the immune response in an area leading to the repulsion of the invader. Antigen reactions with IgE are also responsible for atopic allergic reactions (e.g., hives, asthma, hay fever etc.)
IgD is found on the surface of B-lymphocytes and together with monomeric IgM, serves as antigen receptor for the activation of B cell as described previously. IgD is monovalent.
Important Properties of Antibodies
The immune system in mammals is capable of producing a seemingly unlimited number of antibodies that can react with the millions of antigens we potentially encounter in our lifetimes. This genetic diversity is key to the effective protection of our bodies against pathogens that are constantly evolving new antigen molecules.
Does the cell have millions of genes to create all this variability? This was one early theory, but given the small number of genes in the human (about 25,000), it is clear that this cannot be the case. Antibody diversity was difficult to understand until researchers realized that eukaryotic genes are formed by remarkable recombination events.
The essence of the current theory of antibody diversity is that the genes responsible for the synthesis of a particular antibody are not contiguous units, but are assembled from clusters of gene fragments present in regions of the DNA. One section of these gene fragments codes for the constant region of the antibody, while several other sections, present in multiple copies, code for the variable regions. See Figure 16.4. Somatic recombination allows shuffling of these gene segments into numerous gene combinations that encode for the varieties of light chains and heavy chains that make up complete antibodies. This shuffling begins in the B cell germ line and continues as new B cells are formed and differentiate. Each mature B cell therefore has a unique combination of these regions and makes a unique antibody.
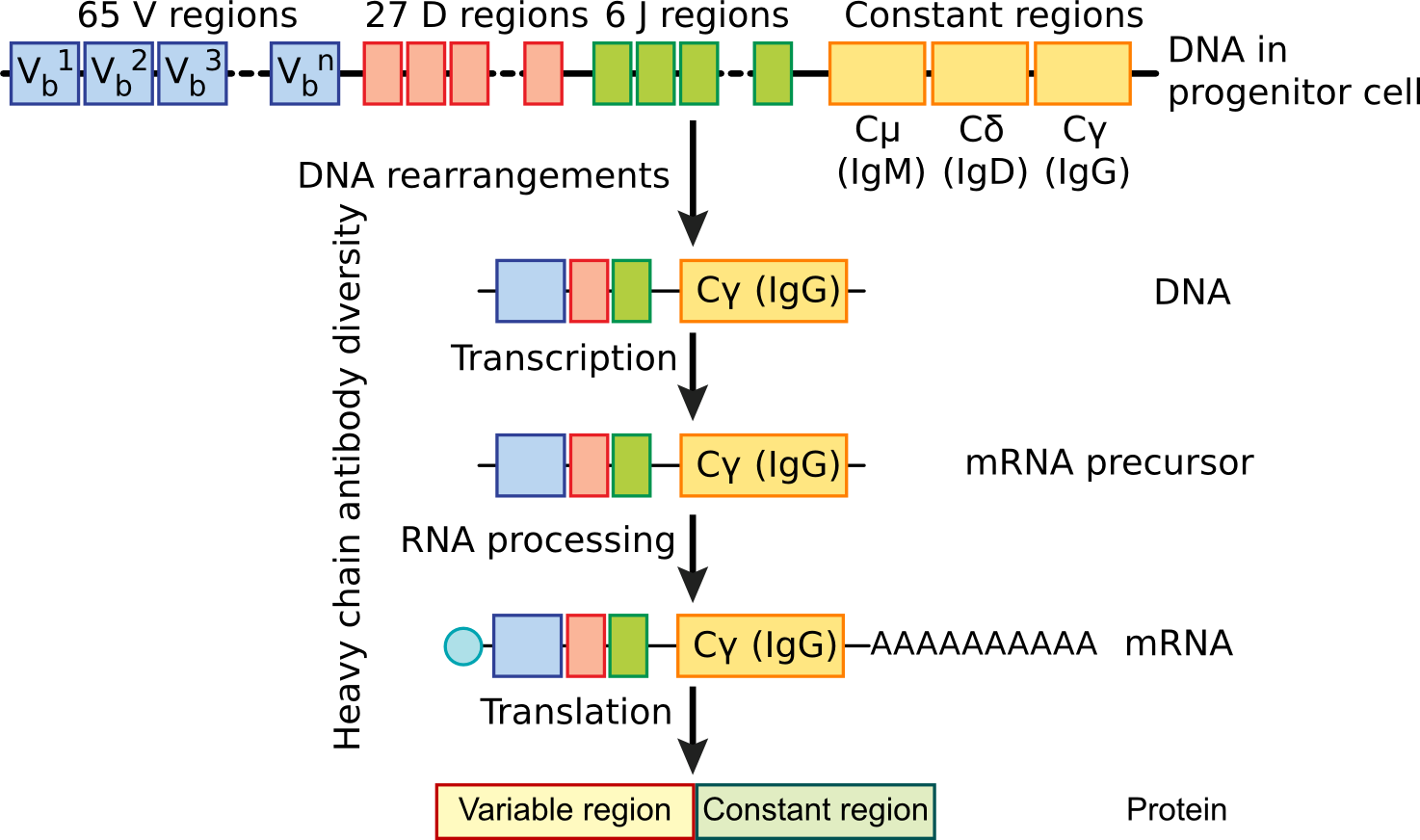
Figure 16.4. B Cell Differentiation. Determination of the variable region of an antibody occurs during the differentiation of the B cell. As it matures, the variable region is randomly picked from several choices and this is spliced to a constant region. Millions of variable regions are possible by this mechanism.
Light chain antibody regions consist of a constant region (C), a joining region (J) and a variable region (V). During the differentiation of the B cell, a deletion joins one V region to one J region. Transcription of the light chain antibody DNA proceeds through the C region and includes the V and J regions. The transcript is subsequently spliced to join the three regions together forming a functional antibody.
Joining of heavy chain antibody genetic regions proceeds in a similar fashion, but is slightly more complex. There are five C regions corresponding to the five classes of antibodies and a third multigenic region, the D (diversity) region, which increases the potential variability of heavy chains. During DNA deletion of the germ line, V, D and J regions are joined together and link to the constant region. Initially, all B cells produce heavy chains with the μ type of constant region that corresponds to IgM antibodies, but later in antibody expression, further splicing can occur causing a different constant region, often the γ region corresponding to IgG, to be produced. This explains why initial antibody production to a new pathogen is high in IgM, but later production shifts to IgG. Also, subsequent infections with the same organism produce more IgG.
Table 16.2. Calculating Antibody Diversity
| Region | Number |
| κ Light chain | |
| V | 40 |
| J | 5 |
| λ Light chain | |
| V | 30 |
| J | 4 |
| 40 x 5 + 30 X 4 = 320 light chain combinations | |
| Heavy chain | |
| V | 65 |
| D | 27 |
| J | 6 |
| 65 x 27 x 6 = 10,530 heavy chain combinations | |
| 320 x 10,530 = 3,368,600 antibody combinations | |
Using the known number of V, D and J regions, it is possible to determine the potential number of different antibodies that could be formed. The actual number is probably more than this as explained in the text.
The amount of antibody diversity that can be generated in the human is diagrammed in Table 16.2. In the germ cell, the DNA in humans contains about 40 V regions and 5 J regions in κ light chains and 30 V regions and 4 J regions in λ light chains. Note that κ light chains and λ light chains are two separate types of light chain constant regions. Assuming that all combinations are possible, that gives a potential diversity of (40 X 5) + (30 X 4) or 320 different light chain combinations. For heavy chains, there are 65 V regions, 27 D regions and 6 J regions for a total of 10,530 heavy chain combinations. Since any light chain can combine with any heavy chain, this give a total of 320 X 10,530 or 3,369,600 possible antibody types.
It turns out that 3 million is actually a dramatic underestimate of the number of actual antibody types because of two additional mechanisms.
- The V region of germ cell line DNA is susceptible to a high rate of mutation resulting in significant amino acid differences even in originally identical V regions.
- Combinational joining between V and J in light chains or V, D and J in heavy chains is deliberately inexact. The junction between these regions can combine at different crossover points, forming different codons. If the number of potential junction points is 10 between each V and J region, then the number of possible antibody types also increases 10-fold.
So the actual number of antibody types is in the hundreds of millions. This large antibody diversity allows reactions with any number of conceivable antigens.
Any pathogen has many different molecules on its cell surface and each one is a potential antigen that the immune system could react to. Each antibody that reacts with a given foreign object has characteristic affinities. Also, for any one antigen, there are usually several different antibodies that react with it and these antibodies can have a whole range of affinities for the antigen. Interestingly, the B cells that have the most reactive and specific antibodies on their surface are stimulated by the antigen to greater extent and end up proliferating more than B cells that react in a weaker fashion. In this way, strongly reacting antibodies are favored and thus the cell can fine-tune its response to an antigen to get the most bang for the buck. This affinity maturation results in the host creating ever more effective antibodies against an infection.
Antibody Reactions
Antibodies bind antigens, but how does this happen at the molecular level? It turns out there are two processes that bring antigens and antibodies together.
- Attractive forces. Attraction between the antigen and the antibody manifests itself in forces we have already talked about in previous chapters. Antibodies and antigens interact by various combinations of H-bonding, electrostatic and hydrophobic interactions, though the exact contribution of each depends on the particular antigen-antibody pair. The combination of these three forces can result in a powerful attraction between an antibody and its antigen, but attraction is not enough.
- Complementary shapes. The shape of the antigen must fit snugly inside the binding site of the antibody as a hand fits inside a glove. Without this tight fit there is weak binding even if there are attractive forces.
Reacting pairs of antigens and antibodies can have very strong binding equilibria, with association constant values as high as 1012, indicating that antibody binding reactions progress a trillion fold faster than the reverse reaction.
Once bound, antibodies use a number of pathways to cause the destruction of the antigen source.
- Complement activation. Antibodies can stimulate the classical pathway of complement activation. This results in cell death due a number of mechanisms as described in Chapter 15.
- Agglutination. Antibodies can cause the agglutination of antigen-bearing entities as shown in Figure 16.5. Remember that every antibody has at least two identical reactive sites. Potentially these sites can bind two antigens. If these happen to be on two different invading pathogens, they will then be stuck together. With the large amounts of antibodies that are produced, the pathogenic cells end up trapped in large clumps and fall out of solution. The host's phagocytes can then easily deal with these networks. IgM antibodies with ten binding sites and IgA with four are especially adept at causing agglutination reactions.
- Toxin neutralization. Antibodies that react with protein toxins almost always cause their inactivation. Toxins play an important role in pathogenesis and their elimination often nullifies the damaging effects of the pathogen. One of the vaccines you receive regularly, TDaP has inactivated diphtheria and tetanus toxins. Being immune to these toxins, makes you immune to these diseases.
- Opsonization. Antibodies are excellent opsonins and greatly accelerate the phagocytosis of pathogens. The constant regions of the IgG antibodies contain binding sites that react specifically with receptors on phagocytes as described previously.
- Prevention of attachment. Antibodies bound to parasites can cover up proteins on the surface of pathogens (e.g., pili on bacteria or hemagglutinin proteins on the outside of the influenza virus) that are necessary for attachment to the target tissue in the host. Masking of these proteins can prevent subsequent infection and secretory IgA probably has a significant amount of its effect in this manner.
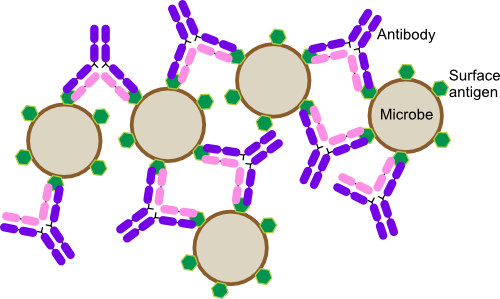
Figure 16.5. Antibody Agglutination. Antibodies have two identical variable regions that can interact with antigen. If the antibody binds to antigens on two separate microbes, it can cause them to stick together. As more antibody attacks the antigen, large clumps of antigen-antibody complexes form and this is called agglutination. Note in this figure the antibodies are drawn much larger than in reality for clarity.
Key Takeaways
- Antibodies are composed of a light chain protein and a heavy chain protein that come together and form a Y-shaped structure. The base of the Y is a conserved region that all antibodies have in common, while the tips of the forks of the Y are unique to each antibody.
- The tips react with the antigen, while the conserved base interacts with the immune system.
- Five types of antibodies are formed in the body: IgG, IgM, IgA, IgD and IgE.
- The variable region of antibodies is formed by spicing together different short segments of DNA during the maturation of the B cell. Using this mechanisms, millions of different variable regions, and thus different antibodies can be made.
- Antibodies bind to antigens by attractive forces and complementary (key-in-lock) shapes.
- Antibodies help defend against pathogens by activating complement, serving as opsonins, agglutinating and sterically hindering pathogens and by neutralizing toxins.
Quickcheck 16-3
Warning, you must be logged in to be able to have your exam graded. Answer the questions below and if you are a registered user of the site you will see a Grade Exam button. Click it to have your exam graded.