Latest News
- Clues beginning to emerge on asymtomatic SARS-CoV-2 infection
- Back in November of 2020, during the first wave of the COVID-19 pandemic, I was teaching an in-person microbiology laboratory. One of my students had just been home to see his parents, and they all c…
- Read more
- Could there maybe be better uses of genetics and probiotics?
- Professor Meng Dong and his laboratory have created a probiotic that can metabolize alcohol quickly and maybe prevent some of the adverse effects of alcohol consumption. The scientists cloned a highl…
- Read more
- ChatGPT is not the end of essays in education
- The takeover of AI is upon us! AI can now take all our jobs, is the click-bait premise you hear from the news. While I cannot predict the future, I am dubious that AI will play such a dubious role in…
- Read more
- Fighting infections with infections
- Multi-drug-resistant bacterial infections are becoming more of an issue, with 1.2 million people dying of previously treatable bacterial infections. Scientists are frantically searching for new metho…
- Read more
- A tale of two colleges
- COVID-19 at the University of Wisconsin this fall has been pretty much a non-issue. While we are wearing masks, full in-person teaching is happening on campus. Bars, restaurants, and all other busine…
- Read more
Chapter 16 - Immunity - Adaptive Immunity and Errors of the Immune System
16 - 1 Introduction
In the previous chapter we introduced the immune system, where it came from, it's primary goal and then explored the innate immunity your body has to fight off infections. These systems are very powerful and effective, but they are immutable and cannot tailor themselves to the threat of an infection. If these defenses are breached, it is up to adaptive immunity to mount a defense.
16 - 2 Adaptive Immunity - introductory concepts
Learning Objectives
- Know that adaptive immunity involves humoral immunity and cell-mediated immunity.
- Describe the role of dendritic cells in the immune system.
- Understand that the immune system is specific, it remembers a previous exposure to an antigen, and it tolerates the presence of its own macromolecular components.
- Describe the actions of B lymphocytes when exposed to antigen.
In contrast to the innate immune system, the adaptive immune system's initial response to a pathogen is weak or non-existent, but as the host has time to respond, this system becomes quite vigorous. To respond to a pathogen, the adaptive immune system must learn to recognize it. The mental picture to have in your mind is that of the body taking bits and pieces of the pathogen (its antigens) and presenting them to certain cells of the adaptive immune system. Those cells that can react with the antigen then marshal a bilateral response with one set of cells producing antibodies (humoral immunity) and the other set activating a group of cells to attack the pathogen (cell-mediated immunity). In this part, we will describe the cells that are involved in the processing and response to antigens, followed by the next section when we will put it all together and look at the reaction of the entire immune system to two different types of pathogens.
In addition to their role in innate immunity, phagocytic cells also play a critical role in adaptive immunity. Macrophages and monocytes take a portion of the debris left over from the digestion of a pathogen and provide that as antigens to the adaptive immune system. This ability is not only present in macrophages but B-lymphocytes and dendritic cells as well. These cells are collectively referred to as antigen presenting cells. One of the more important cell types capable of antigen presentation are dendritic cells.
Dendritic Cells
Most tissues of the body contain a cell type with long tendrils that wrap around and between neighboring cells. Because these branch-like projections are so similar to the intricate dendrites of nerve cells, these immune system components are called dendritic cells. There are four populations of dendritic cells named for their origin or location. Dendritic cells, Figure 16.1, are found in skin, mucous membranes, bloodstream, lymph and all solid organs of the body except the brain, the eyes and testes.
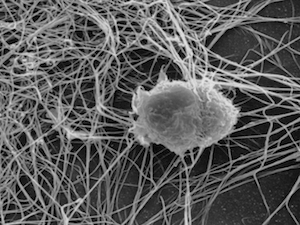
Figure 16.1. A Dendritic Cell. Dendritic cells are found in most tissues of the body, projecting their branches throughout the immediate area, they serve to monitor foreign antigens that may exist. If an antigen if found, it is taken up by the dendritic cell and presented to the immune system. File used with permission, by By Aszakal [CC BY-SA 3.0 or GFDL], via Wikimedia Commons
{kind=link}
These cells play at least three roles in the body.
- Dendritic cells in various tissues play a pivotal role in activating the immune system. Due to their placement throughout the body and their projections snaking through tissues, they are among the first cells to be exposed to invading pathogens. During the response to a parasite they take up antigens from it and present these to the immune system.
- Dendritic cells located in the thymus help to educate immature T cells by exposing them to various antigens (see below).
- Dendritic cells maintain the stimulation of B cells so that they continue to synthesize antibodies. Dendritic cells located in various lymphoid tissues carry out this role.
Dendritic cells are the only cells that can activate a naive T-cell and are the most potent antigen-presenting cells in the body.
Lymphocytes Make Antibodies and Regulate the Immune System
Lymphocytes are the other major type of white blood cells important in the immune system, and they work in concert with phagocytic cells to combat infections. However, the role lymphocytes play in fighting infections is very different than that played by phagocytes. Lymphocytes, with the help of antigen-presenting cells, are part of what is called the adaptive immune system. This system has three important features that all depend on lymphocytes.
- Specificity. B cells/antibodies or T cells react specifically with the antigen that activated them and they generally do not react with any other antigens with a different stucture. These reactions are as specific as the binding of substrate to the active site of an enzyme; in fact, antigen-antibody reactions share many features with enzyme-substrate binding.
- Memory. The immune system remembers an antigen after exposure to it. During the first immunological response to an antigen, lymphocytes capable of responding to that antigen (due to their specific membrane receptors) increase in number and remain present after the antigen is removed. During a second challenge by the same antigen, the immune response of this larger population of lymphocytes produces more antibodies and activates T cells faster, resulting in the swift removal of the entity producing the offending antigen. This type of response is referred to as a memory or secondary response.
- Tolerance. A healthy mammal does not react to its own macromolecular components, all of which are potentially antigenic. The animal is said to be tolerant of its self-antigens. The body achieves this tolerance by eliminating lymphocytes that react to self.
Cytokines and Chemokines
In many cases, the cells in the immune system need to communicate with other cells. When a cell is infected with a virus, it acts to notify its neighbors. When a tissue is damaged by bacterial invasion or physical injury, the surrounding tissue must be informed to begin the repair process. When cells are differentiating in the bone marrow, modulation of gene expression dictates the final cell type. In all of these cases, this communication takes place in the form of small proteins called cytokines and chemokines.
Cytokines are a group of small (less than 30 kDa) soluble proteins that regulate cellular function. Cytokines made by lymphocytes are called lymphokines. Table 16.1 lists some of the more important cytokines that play a role in the immune response. T helper cells, macrophages and monocytes produce many of these cytokines. Some of the most important cytokines are those that communicate between leukocytes and these are given the designation interleukins.
Table 16.1. Important Cytokines and Chemokines in the Immune Response
| Cytokine or Chemokine | Producing cell | Target cell | Effect |
| Interleukin-1 (IL-1) | Macrophages, monocytes and B cells | T cells, B cells | Acts as a growth regulator of T cells and B cells. Induces other cells such as hepatocytes to produce proteins relevant to host defense Serves as an endogenous pyrogen, which produces fever. |
| Interleukin-2 (IL-2) | T cells | T cells | Stimulates the proliferation of T cells and activates natural killer cells. |
| Interleukin-3 (IL-3) | Stem cells, Mast cells | Regulates the proliferation of stem cells and the differentiation of mast cells. | |
| Interleukin-4 (IL-4) | TH2 | B cells | B cell proliferation and enhanced antibody synthesis. |
| Interleukin-5 (IL-5) | TH2 | B cells | B cell differentiation and IgA synthesis |
| Interleukin-6 (IL-6) | TH2, monocytes, macrophages | B cells, plasma cells, stem cells | B cell differentiation and antibody production. T cell activation, growth and differentiation. Has a major role in the mediation of the inflammatory and immune responses initiated by infection or injury. |
| CXCL-8 | Many host cells | T cells, neutrophils, macrophages | Chemoattractant for neutrophils |
| α-Interferon (IFN-α) | Leucocytes, tissue cells | Tissue cells | Inhibition of viruses |
| γ-Interferon (IFN-γ) | T cells | Tissue cells, macrophages, natural killer cells | Inhibition of protein synthesis in virally infected cells. Activation of macrophages and natural killer cells. Stimulates IL-1, IL-2 and antibody production. |
| Tumor Necrosis Factor-α (TNF-α) | T cells | Tissue cells (tumors) | Kills cells, including tumor cells |
| Tumor Necrosis Factor-β (TNF-β) | T cells | Tissue cells (tumors) | Kills cells, including tumor cells |
| Colony Stimulating Factors (CSF) | TH1, macrophages | Phagocytes | Causes phagocytic white cells of all types to differentiate and divide. |
| Macrophage chemoattractant and activating factor (MCAF) | Monocytes, macrophages, fibroblasts (connective tissue cells) and keratinocytes (skin cells) | Macrophages, T cells | Attract and activate macrophages and T cells. |
| B cell growth factors | T cells | B cells | Multiplication of B cells |
A large number of signal molecules are manufactured by various tissues of the body. The cytokines and chemokines listed here are important in various parts of the immune response.
Another important group of signal proteins in immunity are the chemokines. Chemokines are a set of small molecular weight proteins (8-12 kDa) that serve as chemoattractants to phagocytes and T cells. Damage to cellular tissue from bacterial invasion, viral infection or physical injury causes the secretion of chemokines. Since many parts of the body are susceptible to this type of damage, it is not surprising that a large assortment of cell types, including lymphocytes, are capable of synthesizing chemokines. Their production attracts T cells and phagocytes to the area of damage stimulating an inflammatory and immune response.
Key Takeaways
- Adaptive immunity involves humoral immunity and cell-mediated immunity.
- Dendritic cells ingest foreign antigens in the body and present them to the immune cells. They are also important in educating immature T cells and help to maintain the stimulation of B cells so that they continue to make antibodies.
- The immune system has three important properties: It is specific, it remembers a previous exposure to an antigen, and it tolerates the presence of its own macromolecular components.
- Cells of the immune system (and other cells of the body) communicate by the secretion of small (< 30 kDa) proteins called cytokines. Chemokines are 8-12 kDa proteins that serve as chemoattractants to phagocytes and T cells.
16 - 3 B Lymphocytes - Humoral Immunity
Learning Objectives
- Describe the structure of antibodies and the 5 types that are found in the body.
- Describe the mechanisms used by the cell to create millions of antibodies.
- Explain how antibodies defend against pathogens.
B lymphocytes or B cells are the class of immune cells that synthesize antibodies and are responsible for humoral or antibody-mediated immunity. Antibodies react with antigens and the immune system has a virtually unlimited capacity to create antibodies that react with millions of potential antigens.
Each B cell contains on its surface about 100,000 identical antibody molecules (IgD or IgM) The binding sites of these antibodies will all react specifically to the same antigenic structure, but each unique B cell will contain a different antibody, reactive with a different antigen. These membrane-bound antibodies serve as receptors for the antigen and contain two additional polypeptides (dimers of a protein called Ig-α/Ig-β) that span the cellular membrane and play a role in transmitting the binding of antigen to antibody to the cytoplasm of the cell as shown in Figure 16.2. Binding of an antigen by several different antibody receptors on the surface of the cell causes the receptors to cluster in a small area. This clumping of membrane bound antibody molecules then activates a signaling cascade that eventually results in the transcription of genes and thus the production of proteins important for B cell activation.
Figure 16.2. B cell activation. Binding of antibody activates tyrosine kinase and tyrosine phosphorylase in the cytoplasm that then phosphorylates and dephosphorylates tyrosine residues on the Ig-α/Ig-β polypeptide. These phosphorylation/dephosphorylation reactions activate B cells by at least three different pathways: (i) Activated tyrosine residues on the Ig-α/Ig-β polypeptide cause phospholipase C activity to increase. Phospholipase C then cleaves phospholipids into inositol triphosphate and diacyl glycerol. Diacyl glycerol activates protein kinase C eventually leading to the formation of nuclear factor NF-κB. (ii) Inositol triphosphate causes Ca2+ influx from the endoplasmic reticulum and the outside environment. Increased Ca2+ concentrations activate calmodulin that in turn phosphorylates Ets-1, a DNA binding protein. (iii) Activated tyrosine residues on the Ig-α/Ig-β polypeptide activate the p21ras protein, which leads to the activation of a serine/threonine kinase. This in turn phosphorylates cJUN, another DNA binding protein. NF-κB, Ets-1 and cJUN then travel to the nucleus and cause the transcription of specific genes important in B cell activation.
The activated B cell then goes through a process of rapid division (termed clonal expansion) where many more copies of the cell are made. Some of these cells differentiate into plasma cells that produce large amounts of antibody. The antibody molecules produced specifically react with the antigen that caused the initial activation of the B cell. A fraction of the cells from the clonal expansion remain as undifferentiated B memory cells that are capable of reacting with a second antigen challenge at a later time. If a future challenge does appear, this secondary response is much more rapid, normally inactivating the pathogen before a detectable infection appears.
The “B" of B cell comes from the Bursa of Fabricius, where B cells differentiate in birds. (Some of the early research on the immune system was done in chickens). In mammals, B cells mature in the bone marrow and then travel in the bloodstream eventually settling in immune tissues throughout the body with high concentrations in the lymph nodes and spleen.
Antibodies Come in Five Different Classes
We will now spend a bit more time looking at the structure and function of antibodies to give you a more in-depth understanding of their different types and roles in the immune response.
Antibodies have the interesting challenge of needing to respond to a wide array of antigens, yet still be recognizable to the immune system. They therefore, need to interact with two different types of macromolecules, antigens and parts of the immune system. If antibodies were too similar to each other in structure, they would not be able to recognize many antigens, but if they were too different, they would not have a consistent enough structure to react with various immune components. Antibodies solve this dilemma by having two regions, a highly variable region that recognizes the antigens and constant regions that interact with the immune system.
Plasma cells synthesize a number of different types of antibodies and these serve different functions for the cell. Antibodies are proteins made up of two light chains and two heavy chains, see Figure 15.1. The heavy chain determines the type of antibody class and is bound to the light chain by sulfhydryl linkages. Each chain has a constant region that is similar amongst all antibodies. The constant region interacts with the rest of the immune system to facilitate the immune response. Each chain also has a variable region containing a high degree of amino acid variability that is unique to each antibody and this is the part that binds to the antigen. The proteins wrap together and form a Y structure with the two prongs of the Y containing the variable (Fab) regions and the bottom stem of the Y containing the constant (Fc) region.
The different classes of antibodies are defined by the structure of the constant region of the heavy chain. Each heavy chain gives the class of antibody certain properties. Here we first introduce the properties of these antibody classes and then describe the various roles antibodies play in the inactivation of pathogenic microorganisms.
Classes Of Antibodies
While billions of different variable regions are made, the general structure of antibodies falls into just five classes and this is based upon the type of heavy chain present in the antibody as shown in Figure 16.3. Immunoglobulin G (IgG) is the most abundant circulating antibody, making up 80% of the total antibodies and 75% of that found in serum. It contains a single antibody protein complex, with two heavy chains and two light chains. IgG is the second type of antibody synthesized in response to an infection and is the only antibody that can pass through the wall of small blood vessels to access antigens present in the extracellular spaces. It is also the only antibody capable of crossing the placenta in humans, where it confers the mother's immunity onto the fetus and newborn. This immunity protects a baby for the first 6-12 months of its life and allows it time for its own immune system to mature. IgG is particularly effective at attacking extracellular viruses and protein toxins and is also capable of activating the classic pathway of the complement cascade. It helps to prevent the systemic spread of infection and facilitates recovery from many infections. Finally, IgG is the antibody that serves as an efficient handle for phagocytes, allowing phagocytes to bind to a pathogen through Fc receptors on IgG and rapidly phagocytize a pathogen.
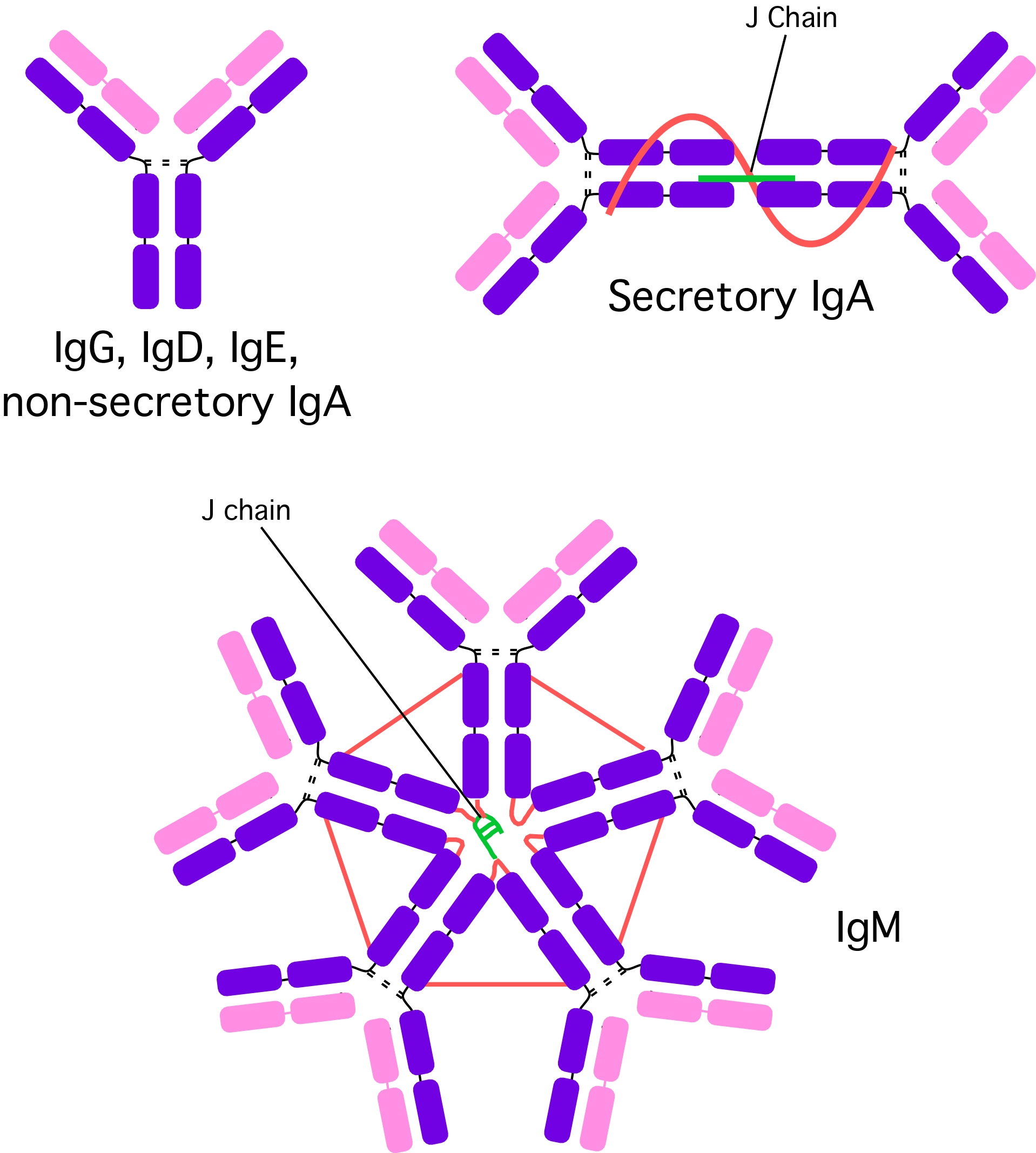
Figure 16.3. Different Antibody Types. There are five different antibodies classes that are formed in the cell. See the text for more details.
IgM is the largest antibody, with five Y structures being joined by their Fc regions in a circular configuration. A J chain (another polypeptide) links the five antibodies together. IgM is present in serum, making up about 10 % of antibodies in the blood. The presence of its ten antigen reactive sites helps agglutinate or clump antigens (see the explanation of this term in the next section), making it easier for the immune system to eliminate them. IgM is more efficient than IgG at activating the complement pathway. IgM is synthesized by plasma cells early in a primary infection and is very important in slowing or stopping the spread of a pathogen during the initial stages of an illness. IgM is also found on mature B cells in a monovalent form, where it serves as a receptor.
IgA is present in serum, mucus, saliva, tears, sweat and milk. Two subclasses with different heavy chains are made, IgA1 and IgA2. IgA1 is synthesized in the bone marrow and makes up most of the serum IgA. IgA2 is synthesized by B cells present in MALT. The antibodies are synthesized as dimers that are joined by a short J chain polypeptide. As the secreted IgA2 passes through the intestinal epithelium, a second secretory protein attaches. Dimerization and binding of the J and secretory proteins make IgA more resistant to proteases present in the environments that it protects. IgA in breast milk interferes with the colonization of the GI tract by harmful microorganisms in the first few months of life. The mother's IgA in the GI tract of newborns keeps these pathogens at low populations, preventing them from causing serious disease, but still allowing the stimulation of the infant's own immune system. The newborn thus develops its own immunity while being partially protected by the mother. IgA molecules do not activate the classical complement pathway, but may activate the alternative complement pathway.
IgE is a monomeric antibody that accounts for only 0.002 % of the total serum antibodies. Almost all IgE is bound to tissue cells, especially mast cells and eosinophils in various parts of the body. Contact of IgE with antigen leads to release of a set of signal molecules from the mast cells, which effectively recruits various agents of the immune response to fight the infection. IgE and MALT serve to detect penetrating pathogens and amplify the immune response in an area leading to the repulsion of the invader. Antigen reactions with IgE are also responsible for atopic allergic reactions (e.g., hives, asthma, hay fever etc.)
IgD is found on the surface of B-lymphocytes and together with monomeric IgM, serves as antigen receptor for the activation of B cell as described previously. IgD is monovalent.
Important Properties of Antibodies
The immune system in mammals is capable of producing a seemingly unlimited number of antibodies that can react with the millions of antigens we potentially encounter in our lifetimes. This genetic diversity is key to the effective protection of our bodies against pathogens that are constantly evolving new antigen molecules.
Does the cell have millions of genes to create all this variability? This was one early theory, but given the small number of genes in the human (about 25,000), it is clear that this cannot be the case. Antibody diversity was difficult to understand until researchers realized that eukaryotic genes are formed by remarkable recombination events.
The essence of the current theory of antibody diversity is that the genes responsible for the synthesis of a particular antibody are not contiguous units, but are assembled from clusters of gene fragments present in regions of the DNA. One section of these gene fragments codes for the constant region of the antibody, while several other sections, present in multiple copies, code for the variable regions. See Figure 16.4. Somatic recombination allows shuffling of these gene segments into numerous gene combinations that encode for the varieties of light chains and heavy chains that make up complete antibodies. This shuffling begins in the B cell germ line and continues as new B cells are formed and differentiate. Each mature B cell therefore has a unique combination of these regions and makes a unique antibody.
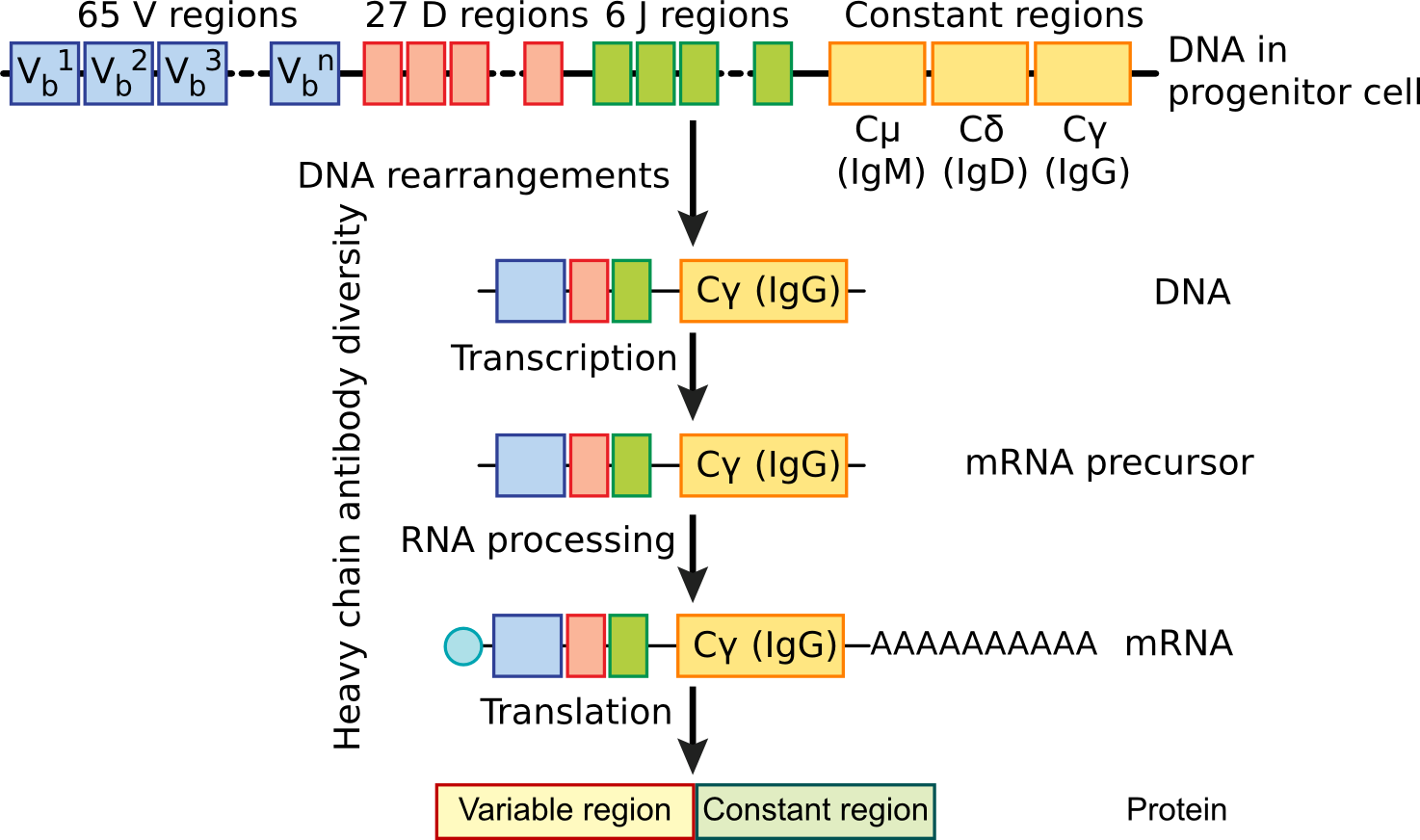
Figure 16.4. B Cell Differentiation. Determination of the variable region of an antibody occurs during the differentiation of the B cell. As it matures, the variable region is randomly picked from several choices and this is spliced to a constant region. Millions of variable regions are possible by this mechanism.
Light chain antibody regions consist of a constant region (C), a joining region (J) and a variable region (V). During the differentiation of the B cell, a deletion joins one V region to one J region. Transcription of the light chain antibody DNA proceeds through the C region and includes the V and J regions. The transcript is subsequently spliced to join the three regions together forming a functional antibody.
Joining of heavy chain antibody genetic regions proceeds in a similar fashion, but is slightly more complex. There are five C regions corresponding to the five classes of antibodies and a third multigenic region, the D (diversity) region, which increases the potential variability of heavy chains. During DNA deletion of the germ line, V, D and J regions are joined together and link to the constant region. Initially, all B cells produce heavy chains with the μ type of constant region that corresponds to IgM antibodies, but later in antibody expression, further splicing can occur causing a different constant region, often the γ region corresponding to IgG, to be produced. This explains why initial antibody production to a new pathogen is high in IgM, but later production shifts to IgG. Also, subsequent infections with the same organism produce more IgG.
Table 16.2. Calculating Antibody Diversity
| Region | Number |
| κ Light chain | |
| V | 40 |
| J | 5 |
| λ Light chain | |
| V | 30 |
| J | 4 |
| 40 x 5 + 30 X 4 = 320 light chain combinations | |
| Heavy chain | |
| V | 65 |
| D | 27 |
| J | 6 |
| 65 x 27 x 6 = 10,530 heavy chain combinations | |
| 320 x 10,530 = 3,368,600 antibody combinations | |
Using the known number of V, D and J regions, it is possible to determine the potential number of different antibodies that could be formed. The actual number is probably more than this as explained in the text.
The amount of antibody diversity that can be generated in the human is diagrammed in Table 16.2. In the germ cell, the DNA in humans contains about 40 V regions and 5 J regions in κ light chains and 30 V regions and 4 J regions in λ light chains. Note that κ light chains and λ light chains are two separate types of light chain constant regions. Assuming that all combinations are possible, that gives a potential diversity of (40 X 5) + (30 X 4) or 320 different light chain combinations. For heavy chains, there are 65 V regions, 27 D regions and 6 J regions for a total of 10,530 heavy chain combinations. Since any light chain can combine with any heavy chain, this give a total of 320 X 10,530 or 3,369,600 possible antibody types.
It turns out that 3 million is actually a dramatic underestimate of the number of actual antibody types because of two additional mechanisms.
- The V region of germ cell line DNA is susceptible to a high rate of mutation resulting in significant amino acid differences even in originally identical V regions.
- Combinational joining between V and J in light chains or V, D and J in heavy chains is deliberately inexact. The junction between these regions can combine at different crossover points, forming different codons. If the number of potential junction points is 10 between each V and J region, then the number of possible antibody types also increases 10-fold.
So the actual number of antibody types is in the hundreds of millions. This large antibody diversity allows reactions with any number of conceivable antigens.
Any pathogen has many different molecules on its cell surface and each one is a potential antigen that the immune system could react to. Each antibody that reacts with a given foreign object has characteristic affinities. Also, for any one antigen, there are usually several different antibodies that react with it and these antibodies can have a whole range of affinities for the antigen. Interestingly, the B cells that have the most reactive and specific antibodies on their surface are stimulated by the antigen to greater extent and end up proliferating more than B cells that react in a weaker fashion. In this way, strongly reacting antibodies are favored and thus the cell can fine-tune its response to an antigen to get the most bang for the buck. This affinity maturation results in the host creating ever more effective antibodies against an infection.
Antibody Reactions
Antibodies bind antigens, but how does this happen at the molecular level? It turns out there are two processes that bring antigens and antibodies together.
- Attractive forces. Attraction between the antigen and the antibody manifests itself in forces we have already talked about in previous chapters. Antibodies and antigens interact by various combinations of H-bonding, electrostatic and hydrophobic interactions, though the exact contribution of each depends on the particular antigen-antibody pair. The combination of these three forces can result in a powerful attraction between an antibody and its antigen, but attraction is not enough.
- Complementary shapes. The shape of the antigen must fit snugly inside the binding site of the antibody as a hand fits inside a glove. Without this tight fit there is weak binding even if there are attractive forces.
Reacting pairs of antigens and antibodies can have very strong binding equilibria, with association constant values as high as 1012, indicating that antibody binding reactions progress a trillion fold faster than the reverse reaction.
Once bound, antibodies use a number of pathways to cause the destruction of the antigen source.
- Complement activation. Antibodies can stimulate the classical pathway of complement activation. This results in cell death due a number of mechanisms as described in Chapter 15.
- Agglutination. Antibodies can cause the agglutination of antigen-bearing entities as shown in Figure 16.5. Remember that every antibody has at least two identical reactive sites. Potentially these sites can bind two antigens. If these happen to be on two different invading pathogens, they will then be stuck together. With the large amounts of antibodies that are produced, the pathogenic cells end up trapped in large clumps and fall out of solution. The host's phagocytes can then easily deal with these networks. IgM antibodies with ten binding sites and IgA with four are especially adept at causing agglutination reactions.
- Toxin neutralization. Antibodies that react with protein toxins almost always cause their inactivation. Toxins play an important role in pathogenesis and their elimination often nullifies the damaging effects of the pathogen. One of the vaccines you receive regularly, TDaP has inactivated diphtheria and tetanus toxins. Being immune to these toxins, makes you immune to these diseases.
- Opsonization. Antibodies are excellent opsonins and greatly accelerate the phagocytosis of pathogens. The constant regions of the IgG antibodies contain binding sites that react specifically with receptors on phagocytes as described previously.
- Prevention of attachment. Antibodies bound to parasites can cover up proteins on the surface of pathogens (e.g., pili on bacteria or hemagglutinin proteins on the outside of the influenza virus) that are necessary for attachment to the target tissue in the host. Masking of these proteins can prevent subsequent infection and secretory IgA probably has a significant amount of its effect in this manner.
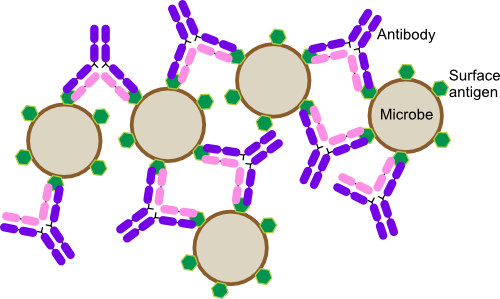
Figure 16.5. Antibody Agglutination. Antibodies have two identical variable regions that can interact with antigen. If the antibody binds to antigens on two separate microbes, it can cause them to stick together. As more antibody attacks the antigen, large clumps of antigen-antibody complexes form and this is called agglutination. Note in this figure the antibodies are drawn much larger than in reality for clarity.
Key Takeaways
- Antibodies are composed of a light chain protein and a heavy chain protein that come together and form a Y-shaped structure. The base of the Y is a conserved region that all antibodies have in common, while the tips of the forks of the Y are unique to each antibody.
- The tips react with the antigen, while the conserved base interacts with the immune system.
- Five types of antibodies are formed in the body: IgG, IgM, IgA, IgD and IgE.
- The variable region of antibodies is formed by spicing together different short segments of DNA during the maturation of the B cell. Using this mechanisms, millions of different variable regions, and thus different antibodies can be made.
- Antibodies bind to antigens by attractive forces and complementary (key-in-lock) shapes.
- Antibodies help defend against pathogens by activating complement, serving as opsonins, agglutinating and sterically hindering pathogens and by neutralizing toxins.
16 - 4 T Lymphocytes Regulate the Immune System and Fight Intracellular Infections, Viral Infection and Cancer
Learning Objectives
- Describe T cells and T cell receptors.
- Explain how T cells interact with antigen presenting cells.
- Describe how T cell mature in the thymus.
- Explain the role of major histocompatibility complex proteins, MHC I and MHC II in the immune response.
- Explain the mechanism of T cell activation.
- Describe the role of cytokines and chemokines.
T lymphocytes or T cells are the second arm of the immune system and are part of what is called cell-mediated immunity. Their roles are to regulate the immune system, help B lymphocytes in the production of antibodies and deal with threats to our body that involve our own cells (e.g., intracellular bacterial infections, viral infections and cancer). T cells recognize antigens in our bodies using receptors on their surface, sensibly called T cell receptors (TCR) as shown in Figure 16.6. These proteins are somewhat reminiscent of antibodies and are composed of two protein fragments, α and β. Each polypeptide chain can be broken into three domains. The carboxy-terminal domain serves as a hydrophobic anchor in the cellular membrane and also contains a disulfide bridge near the outside membrane surface that links the α and β polypeptides. The middle of each protein forms a constant region that shares homology across all TCRs and serves as scaffold for the variable domain projecting furthest into the surrounding environment. TCRs occur in association with CD3 (another protein on the surface of T cells) to form a TCR-CD3 complex. It is this complex that recognizes presented antigens and begins the T cell activation process.
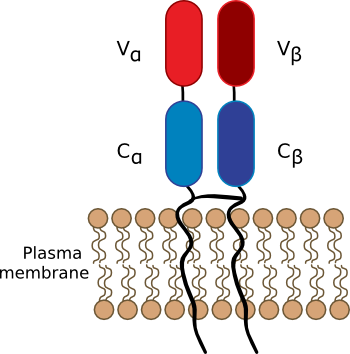
Figure 16.6. The T-Cell Receptor. The structure of the T cell receptor (TCR) is reminiscent of antibody structure. However, T cell receptors are found anchored in the membrane of T cells, while the majority of antibodies float free in various fluids of the body. (Left panel) a cartoon showing the constant and variable regions of the TCR. (Right panel) A molecular model of a TCR. (Source: L. Kjer-Nielsen, et al. 2003. Immunity 18:53–64)
The variable regions of these receptors react with antigens in a fashion that is reminiscent of antigen-antibody reactions. TCRs can respond to a large assortment of molecules and this diversity is achieved by a mechanism similar to that used to create antibody diversity as discussed previously. In contrast to antibodies, TCRs respond to antigens only when it is associated with a membrane protein of another host cell such as antigen-presenting cells (APC). These APCs wrap fragments of foreign molecules inside special proteins, called Major Histocompatibility Complex (MHC) molecules and then display them on their surface for presentation to T cells. In order to understand the mechanism of T cell activation, it is useful to first learn about these important protein complexes present on every animal cell.
Major Histocompatibility Complex Molecules Are the Signal Beacons of the Immune System
Almost all human tissues contain Major Histocompatibility Complex (MHC )molecules, which serve as signal beacons for the immune system. There are two types of MHC molecules. MHC I molecules are found on almost all types of body cells, whereas MHC II molecules are only found on macrophages, dendritic cells and B cells, which can serve as professional antigen-presenting cells. MHC I molecules are status monitors for all cells of the body, while MHC II molecules are important in presenting foreign antigens to other cells of the immune system.
The MHC I molecule consists of a α chain and a β2-microglobulin. One segment of the α chain is anchored to the membrane, with the rest of the molecule protruding outside the cell. Class II MHC molecules are also transmembrane proteins consisting of α and β chains. The two chains fold to give a structure somewhat similar to that of the MHC I molecule, except that both α and β chains are anchored in the membrane. Both MHC I and MHC II molecules contain a deep groove in the domain of the molecule farthest away from the membrane and therefore exposed to the environment. This groove is capable of binding short peptides or other antigens that are not part of the MHC molecule. The peptides in the groove are from various sources, as you will see below. The MHC I groove is closed off and can contain only short peptide sequences, while the MHC II molecule is open and can bind to peptides of longer lengths. Figure 16.7 depicts the structure of MHC proteins.
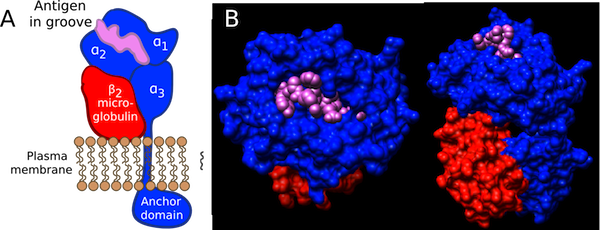
Figure 16.7. Structure of an MHC I Molecule. Major histocompatibility complex proteins are found on most cells of the body. They function to display proteins to T cells. MHC I molecules display proteins that originate in the cytoplasm of the displaying cell. A) A cartoon of the domains of an MHC I molecule. B) Top and side views showing the binding of an antigen (in this case, an influenza peptide) to mouse MHC I molecule. The anchor domain is not shown in the molecular model. (Source: R. Meijers, et al.. 2005. J. Molec. Biol. 345:1099-1110)
Each type of MHC molecule binds peptides from a different source, consistent with their different roles in the immune system. MHC I molecules bind to peptides that originate in the cytoplasm. These peptides were part of cytoplasmic proteins, but were digested by the cellular degradative machinery. This machinery is part of the normal function of a cell and serves to recycle old protein making it available for future use. Some of the short peptide fragments that result from this process are recognized by a specific transporter molecule and translocated from the cytoplasm into the endoplasmic reticulum. Here they are picked up by newly synthesized MHC I molecules and move to the cellular membrane. In this way, the MHC I molecules serve as flags indicating what type of protein processing is going on inside the cell. If the cell is functioning normally, the MHC I molecules contain inside their grooves short peptides derived from normal cell proteins. The immune system recognizes these proteins as self and presumes that all is normal with the cell. In contrast, if the cell is infected by an intracellular parasite, such as virus, some of the MHC I molecules are bound to contain peptide fragments from the virus. These molecules then interact with T cells of the immune system and initiate a response.
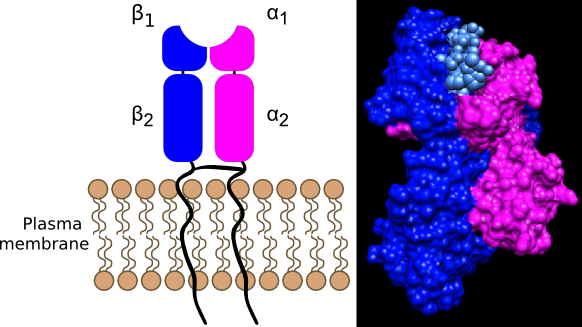
Figure 16.8. An MHC II molecule. MHC II molecules found only on antigen-presenting cells, display foreign antigens picked up in the environment. They contain an α and a β domain. The α1 and β1 domains anchor the protein to the membrane while the α2 and β2 domains form the groove where antigens bind. The MHC II groove is open allowing for the insertion of a larger fragment, 15 to 24 amino acids long. The left panel shows a cartoon of a MHC II molecule, while the right shows the human MHC II molecule binding an antigen (Source Li et. al (2010) J.Mol.Biol. 399: 596-603)
MHC II molecules are present on cells that are part of the immune system and serve as vessels to present foreign antigens to T cells. The structure of the MHC II molecule is shown in Figure 16.8. MHC II molecules bind to antigens that originate from an exogenous source, for example, the leftovers of a phagolysosome after a microorganism has been killed and degraded. Peptide fragments coming from the phagolysosome are combined with preformed MHC II molecules and move to the cell surface. The MHC II molecule, together with the antigen, is then recognized by T cells of the immune system.
There Are Several Types of T Cells, but All Mature in the Thymus
The response T cells have to the presentation of antigens by the MHC molecules on other cells depends upon the type of T cell. Two major subpopulations of T cells differ from one another by the type of surface protein that they display. Mature T cells contain either the CD4 protein or the CD8 protein on their surface. Cells that contain CD4 are further subdivided into four subsets: TH0 cells; T helper 1 cells (TH1); T helper 2 cells (TH2); and pathogenic T cells (T17);. When TH0 cells encounter an antigen they react with, they differentiate into TH1, TH2 or T17 cells, depending the cytokines in the environment, which in turn depends upon the nature of the antigen. TH1 cells participate in cell-mediated immunity and are responsible for recruiting and regulating nonspecific immune cells such as phagocytes. They also help to regulate the steps of inflammation and thus are called T inflammatory cells. TH2 cells serve to activate B cells to differentiate and produce antibody. In most cases, only small amounts of antibody are produced by B cells on their own and TH2 cells are necessary to get full activation of the B cells and large amounts of antibody production. Not much is known about T17 cells, but it is know that their activation increases the severity of some autoimmune diseases (inflammatory bowel disease, multiple sclerosis and rheumatoid arthritis)
The second group of T cells, those with CD8 on their surface, are called T cytotoxic (Tc) cells, also known as cytotoxic T lymphocytes. Mature Tc cells cannot kill target cells until they are activated. This is a two step process.
- The Tc cell has to recognize an antigen presented by a MHC I molecule on a host cell
- The Tc cell has to then be exposed to cytokines (especially IL-2 and IFN-gamma) produced by TH1 cells.
Subsequent presentation of a recognized antigen by a host cell to a Tc cell causes it to attack and kill that cell. The killing involves two mechanisms.
- Tc cells secrete Fas and tumor necrosis factor (TNF) ligands on their surfaces that bind to Fas and TNF receptors on the surface of the target cell. This precipitates a signaling pathway that leads to apoptosis (cell death).
- Fully differentiated Tc cells have granules inside them containing perforin and granzymes. The Tc cell makes contact with the target cell and then releases the perforin. This polymerizes and forms pores in the target cell membrane. The granzymes then release from the Tc cell and migrate through the perforin pores, into the target cell. Once there, the granzymes (serine proteases) activate nucleases and caspases in the target cell resulting in apoptosis.
Tc cells can be thought of as sentries that follow prescribed routes, patrolling our bodies looking for cells displaying odd antigens in their MHC molecules. They also respond to cells that raise the alarm, sent out in the form of an inflammatory response. When found, Tc cells destroy these cells.
T Cell Maturation and Tolerance
T cells originate from bone marrow stem cells in an immature form. Immature T cells can collectively attack a wide range of antigens, including antigens on our own cells. These cells must be selected to only react against non-self antigens and this education takes place in the thymus as shown in Figure 16.9.
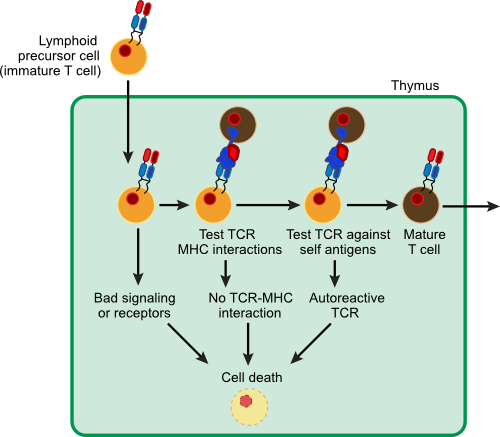
Figure 16.9. T Cell Maturation. Immature T cells move to the thymus for their maturation. The maturation process occurs in two steps. There is a positive selection to ensure that the T cell can interact with MHC molecules correctly. Then there is a negative selection to eliminate those T cells that interact with MHC molecules containing self antigens. T cells that survive this process (only 10%) mature and migrate out of the thymus.
At this point, the education of the T cells begins. Remarkably, immature T cells are initially programmed to die and only through intervention by the thymus do they survive. During their time in the thymus, T cells are exposed to various self-antigens. If they react to the antigen, the T cell is encouraged to carry out its cellular program and die. In fact, the thymus is something of a graveyard for T cells with only about 10% surviving the treatment (some education system!). It may seem more logical to set up the system so that the thymus kills bad T cells, but if you think about it, preprogrammed death is actually safer. If a damaging immature T cell somehow escapes education, it will still die since an educating cell did not tell it to live. In this fashion, the body averts many potential autoimmune reactions. Those T cells that do survive are assumed to be protective against non-self antigens and are released into the bloodstream. These mature T cells then roam throughout the body to fulfill their functions.
Note that B cells undergo a similar type of culling process as T cells to remove self-reactive antibodies, but this process takes place in the bone marrow and not in the thymus. Since the process is very similar to that described for T cells, we will not cover it in detail. Briefly, developing B cells are presented with self-antigens. Reaction between the B cell receptor (remember this is IgD or IgM on the surface of the B cell) and the antigen causes the B cell to die. In this way, the body only produces antibodies to foreign antigens.
The above mechanism is one way the body creates tolerance to self-antigens, but the process is not foolproof. The thymus does not have all the self-antigens of the body available to it and rarely a T cell matures that can potentially react with various parts of the body. In these cases, other mechanisms cope with the presence of a self-reactive T cell, including clonal anergy, active suppression and sequestering.
Clonal anergy is a mechanism where T cells are made unresponsive to antigens, but are not killed. This situation is brought about by T cells reacting with antigen presented on MHC I molecules, but not receiving other signals from the immune system to multiply. Activation of T cells requires presentation of the antigen in the context of a MHC II molecule and a second receptor only found on antigen-presenting cells. Since these are self-antigens, they are not presented to the T cells by an antigen-presenting cell and are only detected on MHC I molecules. The absence of this second signal causes the T cell to shut down and become incapable of releasing IL-2 even during subsequent exposure to the antigen. Since cells expressing MHC I molecules, but not the second signal, are present throughout the body, many self-antigens are readily encountered by a reactive T cell and it is quickly made unresponsive.
Certain T cells, called T suppressor cells, are necessary for turning down the immune system after the source of an antigen has been dealt with. Once a viral or bacterial infection is eliminated, the immune system must be suppressed to prevent further tissue damage. Remember that many of the immune reactions are themselves very damaging to tissue and it is imperative to stop these reactions as soon as they have fulfilled their purpose. T suppressor cells also have a role in protection from self-reactive antigens by mechanisms termed active suppression. It is known that normally healthy humans contain T cells that can potentially cause autoimmune disease and these cells are prevented from causing harm by the action of T suppressor cells. It has also been suggested that T suppressor cells can be induced by reaction with specific antigen, in this case a self-antigen, so that the suppression response matches any unwanted autoimmune reaction of other cells. The mechanism of how this whole process works is still not clear.
A final mechanism to deal with potential autoimmunity is the sequestering (hiding) of potentially stimulatory antigens. Some sites of the body are considered off-limits to the immune system either because the immune system cannot reach the site or for some reason, immune cells cannot detect the antigen due to poor presentation. The classic example is the cornea of the eye. There are no blood vessels that run through this structure, so immune cells have no access to its antigens and therefore, no immune response can be raised. Thus, it is possible to transplant the cornea without fearing rejection by the recipient's immune system. In other cases, certain antigens are hidden away inside cells so that the immune system does not have access to them. When all of these systems fail, it can result in the manifestations of autoimmune disease and we will discuss these later in the chapter.
T Cell Activation Occurs when an Antigen Is Presented to a T Cell and Reacts with Its T Cell Receptor
T cell activation, Figure 16.10, follows a similar pathway to that seen for B cell activation. The best-known activation pathway is that of IL-2 synthesis by TH1 cells and we will go through it in some detail to give you an idea of the molecular mechanisms involved in the process. Similar pathways are present for the activation of other cytokines.
Figure 16.10. Activation of Il-2 Expression in a Helper T Cell. If the TCR recognizes the antigen, a complex is formed between the MHC II molecule, the TCR and the CD4 protein. Successful docking activates a tyrosine kinase in the cytoplasm and this begins a series of cellular reactions. Tyrosine kinase phosphorylates a tyrosine residue on phospholipase Cg1 causing it to become active and split phosphatidyl inositol bisphosphate into inositol trisphosphate and diacylglycerol. Inositol trisphosphate causes a calcium channel to open in the cell membrane resulting in an influx of calcium. The increased calcium levels cause the activation of calmodulin, calcineurin and eventually, nuclear factor of activated T-helper cells (NF-AT). Diacylglycerol activates protein kinase C, which moves into the nucleus and catalyzes the formation of the AP-1 complex from the JUN and POS proteins. NF-AT and AP-1 then form a complex (a transcription factor) that can recognize and bind to a DNA sequence near the IL-2 gene. A second signal is also necessary to activate the T cell. Antigen-presenting cells contain the protein B7-2 on their membranes, which reacts with the CD28 receptor on the T cell. Binding of B7-2 to CD28 activates a separate tyrosine kinase that phosphorylates CD28RC. This protein binds to a site on the IL-2 gene enhancing transcription and also stabilizes IL-2 mRNA once it is formed, increasing its functional concentration. This secondary signal greatly magnifies IL-2 mRNA expression. IL-2 mRNA then moves out of the nucleus for translation at the ribosome.
An antigen-presenting cell that contains a foreign antigen in its MHC II molecule first comes into cell-to-cell contact with a T helper cell and its TCR. Several signals are necessary to activate a T cell. Close contact between the helper T cell and the antigen-presenting cell is required. The first signal is the complex between the MHC II molecule and the TCR, causing the eventual formation of a DNA binding protein (NF-AT/AP-1) that enhances IL-2 transcription. The second signal is between B7-2, another membrane protein on the antigen-presenting cell and the CD28 receptor. This creates a separate signal that results in the formation of a second effector protein, CD28. This then binds to the DNA near the IL-2 gene and also stabilizes the IL-2 mRNA. Both signals, that from the MHC II molecule and the B7-2 protein, are necessary to fully activate the T helper cell.
Another important group of signal proteins in immunity are the chemokines. Chemokines are a set of small molecular weight proteins (8-12 kDa) that serve as chemoattractants to phagocytes and T cells. Damage to cellular tissue from bacterial invasion, viral infection or physical injury causes the secretion of chemokines. Since many parts of the body are susceptible to this type of damage, it is not surprising that a large assortment of cell types, including lymphocytes, are capable of synthesizing chemokines. Their production attracts T cells and phagocytes to the area of damage stimulating an inflammatory and immune response.
A key feature of the adaptive immune response is its memory. After encountering a pathogen and successfully fighting it off, T- and B-memory cells are distributed throughout the body. Upon subsequent challenge by the same pathogen having similar antigens, a much stronger and more rapid response occurs. The enhanced response to a subsequent infection is called a secondary response. It stops the pathogen before it can cause disease. The secondary response is the reason immunity to a pathogen develops. Figure 16.11 shows antibody synthesis during a primary and secondary response.
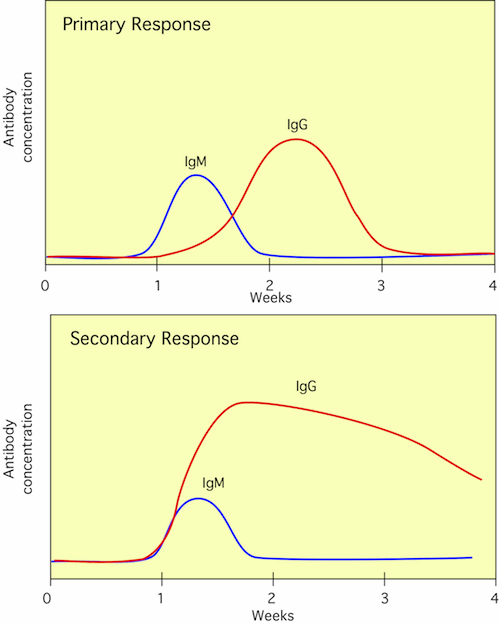
Figure 16.11. A Graph of Antibody Responses IgM and IgG Titers with 1° and 2° Responses to Antigen.. In a first response to antigen (primary response), the antibodies appear more slowly, at a lower concentration, and contain a higher proportion of IgM. During a second exposure to antigen (secondary response) the antibody appears more quickly, reaches a higher concentration and contains more Ig.
Natural Killer Cell Activation Involves Interferon or Il-2, and IgG
We now again return to natural killer cells. Natural killer cells cannot be induced as cytotoxic T cells are but they can be activated by the mechanism of Antibody-Dependent Cell-Mediated Cytotoxicity, also know as ADCC. The mechanism of natural killer attack involves several separate activation steps. Interferon from virally infected cells or IL-2 from T cells must first activate natural killer cells. The second activation step occurs when the natural killer cells encounters a target cell coated with IgG and docks with its Fc region. Having several activation steps prevents the natural killer cell from mistakenly attacking normal cells. Docking then causes changes in the location of cytoplasmic granules and Golgi apparatus within the natural killer cell to a location close to the edge of the cytoplasmic membrane and near the target cell. At this point, the natural killer cell is activated and ready to attack its target. In a mechanisms very similar to that for Tc cells, a pore-forming protein, perforin 1 is inserted into the membrane of the target and the natural killer cell then exports lysosomal secretions into the target cell. The pore made in the membrane together with the attack of the lysosomal contents eventually causes the target cell to lyse and die. The natural killer cell then prepares its cytoplasmic components for the next victim.
Key Takeaways
- T cells respond to antigens using a T cell receptor that is similar in structure to antibodies. Antigen presenting cells present antigen to T cells inside a major histocompatibility complex protein. MHC molecules are membrane-bound proteins: MHC I molecules are found on almost all tissues of the body, while MHC II molecules are found only on antigen-presenting cells.
- MHC molecules possess a deep groove that is capable of holding a short peptide. MHC I molecules process proteins present inside the cell and present them on their surface. MHC II molecules present antigens taken from the phagosome digestion, most often foreign cells, and present them to the immune system. The immune system monitors the proteins present on MHC I molecules and activates when a foreign protein, from an intracellular parasite, is detected. This normally results in the destruction of the cell.
- Mature T cells are classified by their role the immune system. T helper cells help regulate the immune system. Cytotoxic T cells attack cells that have been infected or are displaying odd antigens. In the thymus, T cells are exposed to various self-antigens. If they react, the T cell is allowed to die, if they do not, then the T cell survives. Not all host-reactive T cells are eliminated by their education in the thymus, but these self-reactive T cells are kept under control by clonal anergy, active suppression and sequestering.
- T cell activation occurs when an antigen the T cell reacts to is presented to it in the context of a MHC II molecule. As one example in TH1 cells, binding of T cell receptor to a MHC II molecule causes the formation of proteins that activate DNA transcription of the IL2 gene. IL2 is one example of a cytokine.
- Natural killer cells are activated when a chemokine (either interferon or IL-2) is present and IgG is recognized on the surface of a cell. This causes the natural killer cell to attack, forming a pore in the target cell and pumping lysosomal contents into it causing its death.
16 - 5 Putting It All Together - Examples of Responses of the Immune System to Different Types of Pathogens
Learning Objectives
- Be able to describe how the entire immune system responds to the attack of a bacterial pathogen.
- Be able to describe how the entire immune system responds to the attack of an intracellular parasite.
Chapter 15 and the previous sections in this chapter have introduced the various parts of the immune system and their reactions, but it is helpful to rethink their functions in the context of a complete response to an invading pathogen. In this section, we will examine how the immune system as a whole recognizes and repels an invader and in the process give you a better look at the forest of host defense and not just the trees.
Microbial Pathogens
In this example, we will examine the response of the immune system to Streptococcus pyogenes, a common pathogen of humans. S. pyogenes is responsible for strep throat, scarlet fever, impetigo and (rarely) severe invasive infections, the latter earning it the name flesh-eating bacteria from the media. We will assume that the host has not been previously exposed to this strain of S. pyogenes. Here we will examine one hypothetical scenario, but it is by no means the only outcome of infection with this organism. The process of infection with S. pyogenes and the body's immune response is summarized in Figure 16.12.
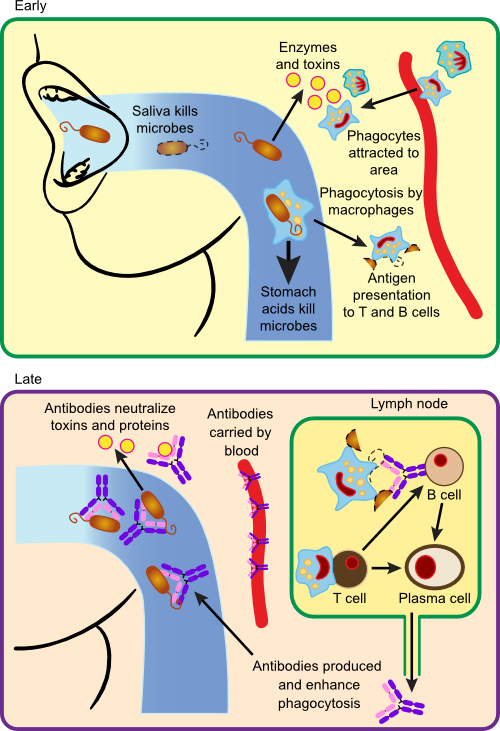
Figure 16.12. Immune Response to Bacterial Infection with S. pyogenes.. Examples of the range of immune defenses against a bacterial pathogen include antibodies and phagocytes that capture and kill the invading microbe. Most of the destruction occurs in the extracellular fluid.
Infection by S. pyogenes typically begins in the mouth and upper respiratory tract. In this example imagine that a group of 10,000 or so S. pyogenes organisms enter the mouth from a droplet of mucus picked up from a nearby child ill with strep throat. Many of them are destroyed by the stomach juices when swallowed and still others are digested by lysozyme in saliva that degrades their outer peptidoglycan layer. The microbes that are left attempt to attach to the surface of the throat using lipoteichoic acid present in their cell walls, but the normal flora occupy most sites for attachment. Many are also repelled by secretory IgA antibody in saliva that cross-reacts with antigens on their surface. However, a few are successful in gaining a loose attachment in the throat and they then bind more tightly with the help of M protein on the microbe and fibronectin present on the epithelial cell surface. Once bound, the microbes begin to multiply and increase in number. Multiplication is hampered by the low pH created by the normal flora and the presence of β-lysins. Lysozyme is also constantly chewing at the cell walls of attached bacteria, but the presence of the capsule protects them from some of these attacks. The colony begins to expand and invade layers below the epithelial cells. The host's surface defenses have failed to repel the infection.
S. pyogenes begins secreting a damaging cocktail of enzymes into the surrounding host tissue. Hyaluronidase degrades hyaluronic acid, a compound that cements cells together in many mammalian systems, allowing further invasion. Secretion of DNase, RNAse and proteases also aid the microbe. This internal invasion causes the secretion of bradykinin and other chemokines from damaged cells, thus initiating the process of inflammation. Bradykinin binds to receptors on blood vessel cells in nearby capillaries causing them to create gaps in the blood vessel walls. Microbial products and the chemokines from damaged cells leak into the surrounding area and into the bloodstream, attracting the attention of phagocytes (mostly neutrophils, but also macrophages) and they begin to migrate to the area and attack the invading S. pyogenes.
At this point phagocytic attack is inefficient, because the bacterium secretes several proteins such as leukocidins, that are toxic to phagocytes. S. pyogenes is also surrounded by a capsule made of hyaluronic acid, yes, the same compound cementing our cell, and this disguise makes the microbes difficult to recognize. Interestingly, this capsule is also attacked by hyaluronidase made by the pathogen, but they seem to make enough of it that it is still protective. The capsule also makes phagocytosis difficult. Despite the resistance of S. pyogenes, a significant number are phagocytized and killed by neutrophils and macrophages. However, due to the slow rate of phagocytosis, the number of bacteria continues to increase. The body attempts to wall off the infection by encasing it in a fibrin clot, but streptokinase produced by the microbe dissolves the clot, further advancing the infection.
As the number of microbes increases, greater numbers of neutrophils penetrate the area. Antigens are also picked up by macrophages and dendritic cells. Active phagocytosis causes the release of more mediators of inflammation and the inflammatory response intensifies. This now becomes noticeable as soreness at the back of the throat. As neutrophils fill with bacteria, they die and make up the pus that runs down the back of the throat and causes the characteristic yellow spots of strep throat.
Bacteria taken up and destroyed by macrophages and dendritic cells are processed and protein fragments, coupled with MHC II molecules are displayed on the cell membrane. During this processing, the macrophages and dendritic cells move toward a nearby lymph node. Inside the lymph node, processed antigens lying in the cleft of the MHC II molecule are presented to a series of T helper cells. Eventually, a successful interaction between the TCR of a TH2 cell and the antigen-presenting cell causes the release of IL-1 and IL-6 from the macrophage. One result of the increased levels of IL-1 is its interaction with the hypothalamus in the brain, which then increases the temperature of the body. The characteristic fever of strep throat increases the activity of T cells. IL-1 and IL-6 stimulate the T cells to proliferate. Proliferating T cells produce IL-2 and IL-4-6 and these further induce T cell multiplication. Most of the TH2 cells help B cells produce antibody, but some remain as memory cells and circulate through the blood and lymph systems.
Receptors on B cells begin to encounter antigens that are flowing from the infected area into the lymph node and these reactions also cause them to proliferate. Some of the antigen that binds to the B cell ends up in MHC II molecules on the surface of the B cell and is presented to T cells residing in the lymph nodes. A previously activated TH2 cell recognizes this antigen in the MHC II molecule and secretes IL-1, IL-4 and IL-5 that stimulate B cells to multiply. As the number of B cells increases, TH2 cells start secreting IL-2, IL-4 and IL-6 that signal to the B cells to differentiate into plasma cells. Plasma cells mobilize large numbers of structures needed for the production and secretion of antibodies (e.g., ribosomes, endoplasmic reticulum, Golgi apparatus etc.). Mature plasma cells migrate to the center of the lymph node and begin secreting IgM antibody specific to the antigen. The rate of secretion can exceed 10 million antibody molecules per minute from an active plasma cell. A fraction of the B cells do not differentiate, but remain as B memory cells. These memory cells disseminate throughout the body in lymphoid tissues and serve as sentries waiting for the next invasion by S. pyogenes.
The full process of antibody induction takes around 7 to 14 days. Initially IgM is made, but this shifts to IgG later in infection. Antibody now exits by the efferent duct, travels through the lymph system and enters into the bloodstream. It circulates in the body and eventually comes in contact with S. pyogenes in the throat.
The entrance of antibody into the sites of infection turns the tide in favor of the host. A large assortment of antibodies is made to many different antigens present on the microorganism. Antibodies directed against the toxins of the microorganism react with and neutralize them. Antibodies against cell surface antigens stick to the microbe and attempt to induce the complement cascade. In the case of S. pyogenes, this reaction is not very effective due to the thick capsule surrounding the microbe. However, antibodies directed against the M protein are extremely effective opsonins. Phagocytes can now rapidly engulf and phagocytize S. pyogenes. Entrance into the phagocyte is rapidly lethal for S. pyogenes, because the microbe does not produce catalase or significant levels of superoxide dismutase and is ill-equipped to deal with the oxygen radicals produced in the phagolysosome. Minutes after being engulfed by a phagocyte, the cells are dead. The end result of antibody production is a rapid clearing of pathogens in the area of infection. Microbial numbers begin to decrease rapidly and within a few days, the infection and its symptoms dissipate.
In reality, this course of events does not happen in a patient with strep throat. Individuals who are infected with S. pyogenes are given penicillin (or another effective antibiotic), to which the microbe has never become resistant. This is fortunate because some of the antigens present on S. pyogenes are similar to self-antigens on the heart valve and cartilage. In about 1-3% of cases, the development of an immune response against S. pyogenes can lead to rheumatic fever, an autoimmune disease where antibodies directed against the M protein of S. pyogenes attack the heart values and joints. To prevent this serious and undesirable immune reaction, S. pyogenes infections are treated with antibiotics before the body has a chance to mount a vigorous immune response.
Host Response to Viral Pathogens Relies Heavily on T Cells
The response to a viral infection is quite different from that seen in a bacterial infection. Viral infections are intracellular for the most part, while the case we described above was extracellular. This has two implications. First, much of the damage is going to occur inside infected cells. Second, elimination of the infection is going to require destruction of host cells. Therefore, the immune system needs a method of discriminating virally infected cells from healthy cells. Fortunately our bodies have just such a mechanism and it involves the MHC I molecules. In general, cell-mediated immunity is much more important for clearing a viral infection. Figure 16.13 summarizes the hosts response to infection with influenza virus.
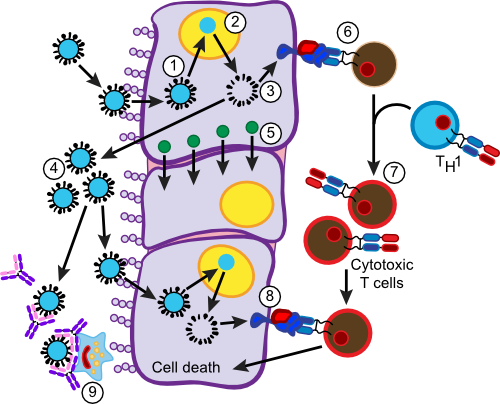
Figure 16.13. Immune Response to Influenza Infection. An immune defense against a viral infection is more dependent on T cells and less dependent on antibodies. Cytotoxic T cells are important in killing virally infected cells. In step (1) influenza virus enters the cell and begins to replicate (2). Viral proteins fill the cytoplasm (3). Most go on to form influenza virus and escape (4). The presence of viral proteins in the cytoplasm also causes the production of α-interferon (5). Some viral proteins are degraded and end up being displayed in MHC I molecules (6). Passing Tc cells will test the MHC I molecule presenting foreign antigen and a fraction will be activated by it. Activated Tc cells are directed to differentiate into cytotoxic T cells by TH1 cells. Activated cytotoxic T cells then attack other virally infected cells displaying viral antigens in the MHC I molecules that the T cells react to. Viral antigens are also presented to B cells producing antibodies in a similar fashion to that described in Section 16.25. These antibodies attack free virus, agglutinating it and making it available for phagocytosis (9).
Influenza virus is a good example to study because it illustrates many of the salient points of viral infection and is also an important human pathogen. However, remember that the specific response of the immune system is dependent upon the particular pathogen. Many of the mechanisms that we describe here come into play in other viral infections, but the exact response is always unique to the particular viral agent.
Influenza is a very contagious disease that easily spreads through respiratory droplets expelled by infected individuals. For this example, imagine that an influenza sufferer has just sneezed on their hand and then opened a door, contaminating the door handle. Another person touching the handle can picked up droplets containing 100,000,000 flu viruses and a subsequent touch can transfer 1,000,000 to the nasal or oral cavity. Some of them then land at the back of the throat. Again, the low pH and unfriendly environment created by the normal flora and host proteins cause the vast majority of the viruses to be inactivated. Those that do survive bind to sialic acid-containing proteins or lipids on the surface of throat epithelial cells and enter by receptor-mediated endocytosis. A drop in the pH of the endosome causes a conformational change in the virus and the release of the eight genomic fragments of influenza virus into the cell. Up until this point, the immune system has no indication that anything is amiss.
Virus begins to replicate and viral proteins accumulate in the infected cell. Some of these proteins are degraded by host cell machinery and the peptide fragments are transported into the endoplasmic reticulum, where they combine with newly synthesized MHC I molecules. The MHC I molecules loaded with foreign viral antigens now find their way to the surface of the cell. The infected cell also begins to produce and secrete α-interferon. This notifies surrounding cells of viral infection and induces them to produce compounds that interfere with viral replication making further infection more difficult.
Virally infected cells begin to release flu virus particles into the surrounding tissues. The presence of viral particles and the death of virally infected cells begin the process of inflammation as described for S. pyogenes infection. This causes the characteristic redness, soreness and swelling in the back of the throat and the induction of fever observed in influenza. At this point, macrophages and dendritic cells take up viruses and viral debris, process them and add to MHC II molecules for presentation to T helper cells. TH1 cells detect the presented antigen and those that match are activated and begin to secrete IL-2, which has several effects on other T and B cells responding to the infection.
Mucous secretion from the intensifying inflammation begins to cause a runny nose and coughing. The release of interferon and IL1 contribute to the aches and fever associated with influenza. As the concentration of virus increases in the body, these symptoms intensify.
Cytotoxic T cells (Tc cells) roaming in the tissues encounter infected cells presenting viral antigens in their MHC I molecules. Those that have TCRs that respond to the antigen are activated to begin clonal expansion and develop into active cytotoxic cells under the influence of the TH1 cells. During the next encounter with a virally infected cell, these cells again recognize the viral antigen being presented in an MHC I molecule using their TCR, but this time, they are activated to attack the cell. The Tc cell binds to the infected cell and begins a destructive cycle. A number of cytokines, including γ-interferon and tumor necrosis factor (TNF), are secreted by the Tc cell. These factors limit viral replication in the target cell and also attract phagocytes to the area. The Tc cells also produce molecules that elicit a form of programmed cell death (apoptosis) in the target cell, essentially telling the cell to kill itself (see section 16.3). Phagocytes that enter the area then clean up any remaining viral debris. As in the case of B cells, some of the Tc cells of the clonal expansion do not differentiate, but remain as memory cells in preparation for the next viral challenge by this viral strain.
Phagocytized virus is also presented to TH2 cells that respond by activating in a manner identical to that described earlier for bacterial infections. Virally infected cells also stimulate B cells that respond by clonal expansion and differentiation into plasma cells, resulting in the formation of antibodies against viral antigens. Antibodies are commonly raised against hemagglutinin and neuraminidase, two proteins on the outer surface of the virus. In most viral infections, antibody is not as important as in bacterial infections, but it does have several consequences. Antibodies bound to virus cause them to agglutinate, precipitate them out of solution and slow their spread through the body. Many effective antibodies block the receptor site of the virus and prevent its attachment to new host cells. These types of antibodies are especially useful in stopping subsequent infections by the same virus. Antibodies attached to virus also assist phagocytes in the efficient uptake of viral particles.
Free virus in the body is eliminated by the action of antibodies and phagocytes and the activated Tc cells destroy any virally infected cells. As the load of virus present in the body decreases, T suppressor cells help the immune response abate. After infection, subsequent attack by this strain of influenza virus is prevented by the action of T and B memory cells.
Key Takeaways
- The response of the immune system to a pathogen is integrated and involves all of the cells and tissues that have been discussed in this chapter.
- This immune response is different, depending upon the type of pathogen encounter, with a major distinction of response occurring based upon whether an infection is extracellular or intracellular.
16 - 6 Errors of the Immune System
Learning Objectives
- Understand the nature of allergies.
- Describe the four classes of allergic reactions.
- Understand the nature of autoimmune disease.
- Explain how autoimmune diseases can be triggered.
The immune system is amazingly good at protecting the body against the numerous pathogens that challenge us, but immunity also makes mistakes that can cause serious health problems. A common mistake begins when the immune system recognizes a foreign protein that does not pose any threat, sometimes raising a violent immune response. We are of course talking about allergic responses. Rarely, the immune system recognizes self-proteins and begins to attack the body's own tissues, setting the stage for autoimmune diseases. In this section, we will examine what is happening in allergic response and autoimmune diseases at the cellular level.
Allergies
Allergies are responses to foreign molecules that are in our bodies. The problem here is that the foreign molecule does not pose any threat and in many cases, our immune systems suffer chronic exposure to these antigens (called allergens in this context) for long periods of time. This results in an exaggerated (and detrimental) immune response that causes tissue damage. The first exposure to allergen activates the immune response, but it is the second and subsequent exposures that result in the symptoms we associate with allergies. In susceptible individuals, these allergens cause sneezing, coughing, itchy and, watery eyes and sometimes hives. Allergic reactions can be immediate or delayed and the rate and types of reaction indicate different immune responses. Peter Gell and Robert Combs developed a system in 1963 to classify these different reactions and this classification was later found to correlate with four different molecular pathways that lead to allergic responses. The four types were sensibly given the names Type I, II, III and IV hypersensitivity.
Anaphylactic Hypersensitivity (Type I)
Type I hypersensitivity or anaphylactic reactions are characterized by an immediate reaction to the offending allergen upon second contact. This is one of the more common types of allergies and there is a hereditary predisposition for anaphylactic reactions. The process itself involves B cells, mast cells and basophils. It appears that these individuals mount a vigorous immune defense against something that is not harmful, whereas the healthy immune system would not react so strongly. Figure 16.14 shows the steps in an anaphylactic reaction.
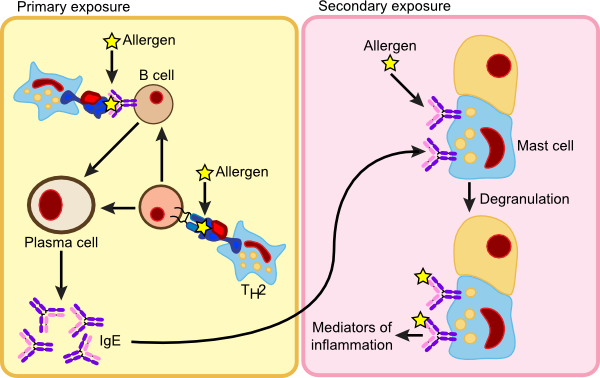
Figure 16.14. An Anaphylactic Reaction. On initial exposure to an allergen, the immune system stimulates B cells that then synthesizes IgE. The IgE migrates through the body and inserts into Mast cells. Upon second exposure, the allergen binds to the IgE, causes the Mast cells to degranulate and dump a collection of substances called the mediators of inflammation. The ensuant reaction can be violent and life-threatening.
During the first exposure to allergen, contact is made with B cells that differentiate and produce IgE antibody. The IgE against the allergen is released into the bloodstream and eventually binds to Fc receptors on mast cells and basophils. The mast cells are now sensitized, lying in wait for the second exposure. Upon contact with allergen a second time, it now attaches to IgE present on mast cells causing degranulation. On the cellular level, the granules present in the cytoplasm migrate to the cellular membrane and spill out their contents into the surrounding area. This results in the release of histamine, slow reacting substance of anaphylaxis (SRS-A), heparin, prostaglandins, platelet-activation factor (PAF), eosinophil chemotactic factor of anaphylaxis and proteolytic enzymes. This cocktail of proteins are the mediators of inflammation and they trigger a number of physiological responses including smooth muscle contraction, an increase in vascular permeability and mucous secretion.
In its most severe form, systemic anaphylaxis, there is a generalized response. Exposure to allergen causes an immediate, large amount of mast cell response over a short period of time. This results in large concentrations of the mediators of inflammation being released all at once. Individuals experiencing systemic anaphylaxis have trouble breathing due to smooth muscle contraction causing the closing of the bronchioles in the lungs. Arterioles also dilate, resulting in a drop in blood pressure and capillary permeability that causes a loss of fluid into tissues. Victims of this response can die within minutes from reduced blood return through the veins, asphyxiation, low blood pressure and circulatory collapse leading to shock. Quick treatment with epinephrine (increases blood flow and inactivates mast cells) and antihistamines (combat histamines) is essential to prevent death. Common allergens in this type of reaction are penicillin, passively administered antisera and insect venom from bees or wasps.
Localized anaphylaxis (atopic allergy) is a less severe form of anaphylaxis, whose symptoms depend primarily upon how the allergen enters the body. In hay fever (allergic rhinitis) the allergen enters the upper respiratory tract. Common allergens in hay fever include pet dander, pollen, fungal spores and household dust mites. Exposure to these particles causes the typical symptoms of hay fever, i.e., runny nose, itchy eyes, coughing and sneezing, most of which are indicative of the action of mast cells. Treatment typically involves the use of antihistamines to block the action of histamine released by mast cells.
Bronchial asthma results when the site of immunological response is the lower respiratory tract. The same allergens that irritate the upper respiratory tract in this case cause the symptoms of asthma. Mucus accumulates in the alveoli (air sacs) of the lungs and smooth muscle contraction of the bronchi narrows the airways and causes the characteristic wheezing of asthma. Bronchodilators that relax the bronchial muscles and expectorants that clear mucous plugs in the lungs can relieve most of the symptoms of asthma, but it is still a serious illness that can be fatal if treatment is delayed for too long.
A third type of localized anaphylaxis can be caused by allergens that enter through the digestive tract. Eruptions of the skin called hives are a strong indication of a true food allergy. Once a food allergy is established, it is usually permanent and the only option is to diminish the reaction with antihistamines or to avoid the suspect food altogether. Wheat, peanuts, soybeans, cow's milk, eggs and less often shellfish are common causes of food allergies.
Skin testing can be done to identify the cause of common allergies. These tests consist of inoculation with small amounts of potential allergens and after a suitable incubation period (usually 24 hours), observation for the appearance of a hard, red welt indicating a positive response. Once the responsible allergen is known, the individual has three choices. The first and most obvious is to avoid the allergen and for most food allergies, this is the preferred choice. For allergens that are difficult or impossible to avoid (e.g., pollen, mold, dust mites etc.), treatment with antihistamines is useful. In severe cases, where the first two choices are unsatisfactory, it is sometimes possible to desensitize an individual by repeated injections with the offending allergen. These subcutaneous injections actually cause the elicitation of a second immune response that raises IgG antibodies to the allergen. The desensitization works because the IgG is able to react with the allergen and remove it before the IgE on the mast cells can react with it. Suppressor T cells may also play a role in desensitization.
Cytotoxic Hypersensitivity (Type II)
Type II hypersensitivity involves the destruction of entire cells. Antigens on the surface of the cells in question stimulate the production of IgM and IgG. The IgM and IgG then bind to the tissue, leading to activation of the complement system through the classical pathway. Complement then precipitates an inflammatory response recruiting neutrophils, eosinophils, macrophages/monocytes, natural killer cells and platelets to the area. This creates an intense immune response that results in the death of the target cells. Cytotoxic responses are immediate and a common example of this type of reaction is when a patient is given the incorrect blood type during a transfusion.
Immune Complex-Mediated (Type III) Hypersensitivity
Immune complex-mediated (Type III) hypersensitivity is characterized by the formation of immune antigen-antibody complexes that accumulate in various tissues. Under normal circumstances, monocytes remove these immune complexes, but in the presence of excess antigen, they overwhelm the body's ability to clear them. The accumulation of these complexes activates the complement cascade and leads to undesirable inflammation and damage to the surrounding tissue. Accumulation of type III hypersensitivity complexes is most commonly observed in the kidneys, blood vessels, the joints and the skin. Figure 16.15 depicts an immune complex hypersensitivity.
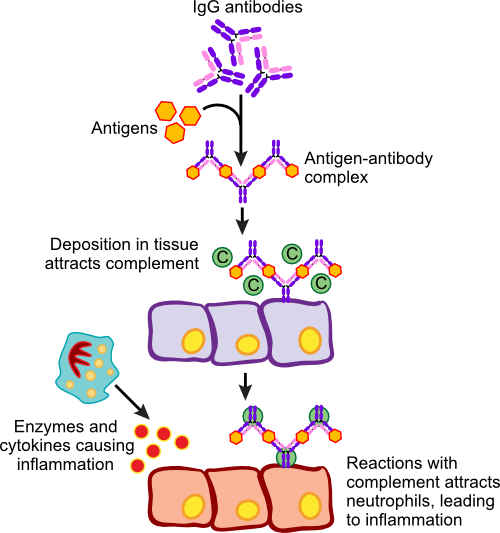
Figure 16.15. Immune Complex-Mediated Hypersensitivity. The chronic exposure to an antigen causes the formation of immune complexes between the antigen and antibodies raised against it. These accumulate in specific areas of the body (kidney, blood vessels, joints and skin) and elicit an inflammatory response that damages the surrounding tissue.
Three types of diseases can lead to immune complex hypersensitivity. First, chronic infection with a virus, bacteria or protozoa, along with a weak antibody response, eventually leads to the deposition of immune complexes (conglomerations of antibody and antigens) and trigger this allergic response. A second type of this condition stems from antibody raised against self-antigens. Since the antigens are not removed in this case, immune complexes are constantly being formed and this is one of the major damaging side effects of autoimmune disease such as systemic lupus erythematosus and rheumatoid arthritis. Third, constant chronic exposure to a particular antigen at a body surface can cause type III hypersensitivity reactions. Farmer's lung is an example of this type of disease. Repeated exposure to fungal spores present in moldy hay elicits an immune response and results in the formation of IgG antibody directed against the spores. Subsequent exposure causes the formation of IgG-spore immune complexes that accumulate in the alveoli of the lungs and cause inflammation leading to lung tissue damage. Other examples of type III hypersensitivity and the allergen that precipitates the problem include, cheese washer's disease (Penicillium casei), Furrier's lung (fox fur protein) and Maple bark stripper's disease (Cryptostroma spores).
Cell-Mediated (Delayed) Hypersensitivity (Type IV)
Cell-mediated or delayed hypersensitivity is related to the T cell response to an allergen. The major factor causing the delay is the time required for T cells that respond to the allergen to migrate to and accumulate at the site of exposure. Delayed hypersensitivity reactions typically begin to appear 24 hours after exposure and reach their maximum intensity in 1 to 3 days. Antigens from microorganisms as well as certain chemicals can cause these types of reactions. Figure 16.16 summarizes the steps in a cell mediated allergic reaction.
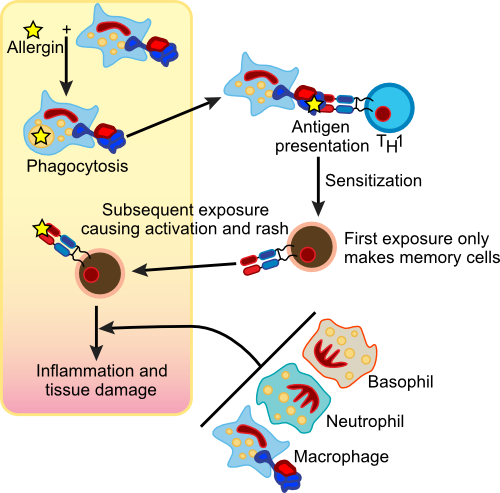
Figure 16.16. Cell-Mediated Hypersensitivity. In this type of allergic reaction, T cells react to the allergen and mobilize the rest of the immune system. An immune reaction, including inflammation, causes the symptoms associated with cell-mediated hypersensitivity and can lead to extensive host tissue damage.
In delayed type hypersensitivity, the allergen is first phagocytized by a macrophage and presented to T cells. The T cells that cause this type of response had been given the name delayed-type hypersensitivity T cells or TDTH cells, but are now know to be TH1 cells. TH1 cells activated by the allergen respond by proliferating, migrate to the area of allergen and then release cytokines, which attract lymphocytes, macrophages and basophils to the tissue. The resulting immune response can cause extensive tissue damage.
Delayed type hypersensitivity reactions are familiar to anyone who has suffered after brushing up against poison ivy or poison oak. In this case, the cause of the reaction is contact with catechols present in the oily leaves of the poison ivy and poison oak plant. Catechols on the skin result in the formation of a characteristic rash with intense itching, swelling and blistering. The catechols are low molecular weight chemicals (haptens) that are capable of combining with a protein to form an antigen. Contact with leaves of the poison ivy plant causes some of the catechols to rub off and stick to lipids in the skin and eventually combine with a skin protein to form the allergen. This then sensitizes the TH1 cells. During initial exposure, only TH1 memory cells are created and no dermatitis occurs. In second and subsequent exposures, the TH1 memory cells become active, recruit other immune cells and cause the characteristic rash. Soaps, cosmetics and metals (especially nickel) can also cause this type of contact dermatitis in sensitive individuals.
Important chronic diseases involve tissue destruction by delayed type hypersensitivity reactions. Certain viruses, mycobacteria, protozoa and fungi are capable of causing these immune responses. In all cases, there is chronic stimulation of T cells leading to significant tissue destruction. Infectious diseases such as leprosy, tuberculosis, leishmaniasis, candidiasis and herpes simplex are associated with type IV hypersensitivity. Delayed type hypersensitivity is the basis for skin tests that detect previous exposure to etiologic agents of tuberculosis, leprosy and histoplasmosis, and other diseases (Figure 16.17).
Figure 16.17. Skin Testing. Pictures of skin testing done to determine the allergens that cause an immune response. Tuberculin allergen is available commercially (A). A small amount of allergen is inject under the skin. If an individual is allergic to a substance an immune response will manifest in the form of redness and swelling (B). The size of the reaction is measured and this is used to gauge a person’s exposure to tuberculosis (C). This individual has been exposed to TB in the past. This is an example of a Type IV allergic response. (Source: Centers for Disease Control and Prevention)
Autoimmune Diseases Involve the Host Immune System Attacking Host Proteins
Under normal conditions, the body is able to distinguish self from non-self antigens. This immune tolerance develops by several mechanisms as described earlier in the chapter. When this tolerance breaks down, autoimmunity occurs, which is characterized by an immune response against antigens present on host tissue. As an individual ages, various types of infections and other encounters with foreign antigens can precipitate the formation of antibodies that react against self-antigens. These are usually benign and under normal circumstances disappear as soon as the offending antigen is removed. Autoimmune disease is often precipitated by the activation of self-reactive T and B cells following stimulation by infection or exposure to certain antigens present in the environment. These illnesses can be extremely debilitating and cause significant suffering in afflicted individuals. Table 16.3 summarizes various autoimmune diseases.
Table 16.3. Various Autoimmune Diseases
| Disease | Tissue affected | Molecular details |
| Type 1 diabetes | Pancreas | Attack of islet cell cytoplasm and surface antigens by cell-mediated immunity and antibodies |
| Male infertility (some types) | Testes | Reaction against surface antigens on spermatozoa by antibodies. |
| Myasthenia gravis | Neuromuscular junction | Antibody against the acetylcholine receptor on the surface of these cells |
| Graves' disease | Thyroid gland | Antibody against thyroid-stimulating hormone receptor |
| Multiple sclerosis | Nerves | Attack of immune system against the myelin sheath of nerves |
| Rheumatoid arthritis | Joints | Reaction of IgG with collagen |
| Idiopathic thrombocytopenic purpura | Platelets | Attack of platelets by antibody resulting in their phagocytosis |
| Systemic lupus erythematosus | Various | Antibodies against DNA, nucleoprotein, cytoplasmic soluble antigen and an array of other antigens. |
| Ulcerative colitis | LPS | Unknown |
A number of autoimmune disease are caused by immune responses to various tissues of the body.
Autoimmune disease is mitigated by four factors.
- There seems to be a genetic predisposition for some autoimmune diseases. For example, there is a correlation between a person's susceptibility to Graves' disease and the presence of a specific antigenic determinant on the MHC molecule. Genes encoding MHC molecules actually have a number of alleles in humans, and as a result, their protein sequence is not the same from person to person.
- Viral or bacterial infections of some sorts can precipitate autoimmune disease. It seems an infection with certain pathogens triggers an immune response that cross-reacts with antigens present in the body. One emerging example is Guillain-Barré syndrome where at least some of the cases appear to be caused by a previous infection with Campylobacter jejuni. After infection, the immune system begins attacking the nervous system causing weakness, sometimes to the point of impeding breathing.
- Endocrine hormones (hormones secreted directly into the blood or lymph) affect the severity of autoimmune disease. Estrogen promotes autoimmune reactions, while androgen inhibits them through yet unknown mechanisms and this partially explains the predominance of autoimmune disorders in women.
- Stress and neurotransmitters (those with effects on the nervous system) can cause physiological changes that make autoimmune diseases worse.
For example, autoimmune diseases cause a progressive degeneration of the tissues attacked by the immune system. In systemic lupus erythematosus, the immune response is errantly directed against a number of different self-antigens including DNA, nucleoproteins and blood-clotting factors. The illness results in the accumulation of antigen-antibody complexes that migrate to the kidneys and joints causing immune complex-mediated hypersensitivity. If untreated, this leads to kidney failure and severe arthritis. Type 1 diabetes is an autoimmune disease, where the immune system responds to the cytoplasm and surface proteins present on islets cells of the pancreas. Destruction of these cells results in the inability to make insulin. Many of these diseases are extremely difficult to treat.
Key Takeaways
- Allergies are caused when the immune system mistakenly reacts to a foreign antigen that is no threat to the body.
- There are four classes of allergic reactions: anaphylactic hypersensitivity, cytotoxic hypersensitivity, immune complex-mediated hypersensitivity and cell-mediated hypersensitivity.
- A second mistake that can be made by the immune system is the recognition of self-antigens. This causes autoimmune diseases that are characterized by constant overstimulation of the immune system.
- Autoimmune diseases are often triggered by infection or exposure to certain antigens in the environment.
16 - 7 Summary
The immune system has three key abilities. First is the ability to detect entities that might do us harm. Innate to this system is the ability to recognize antigens at the molecular level, and antibodies and T cell receptors are the key proteins. Each antibody or T cell receptor recognizes a unique shape and this variability is achieved through the random mixing of genes during the differentiation of T and B cells. Second, the immune system is tolerant. Since antibody and T cell receptor differentiation is a random process, immature B cells and T cells are created that react with our own macromolecules. These potentially lethal cells are identified and nullified by a number of mechanisms. This last part is the key step that allows the immune system to only attack antigens that are non-self. Third, the immune system is adaptive. After an initial challenge by an antigen, the immune system has a memory in the form of the aptly named memory cells. A second challenge by a similar antigen elicits a more rapid and stronger immune response and, in the case of a pathogen, prevents it from causing disease. One great advance in treating disease has been the activation of the immune system against a pathogen by presenting its antigens, but in a form that does not cause disease. We are, of course, referring to vaccination and its use has been one of the major methods of preventing infectious diseases, as we will describe in Chapter 17.
Our bodies are constantly bombarded with trillions of viruses and billions of microorganisms. To withstand this onslaught, multicellular organisms evolved various abilities to defend themselves against these invaders. While these pathogens are resourceful and persistent, the multi-faceted nature of the immune system and its adaptability successfully fends off these attacks. The immune system is critical to our survival and the constantly increasing knowledge that we have of its function will likely lead to many revolutionary treatments in the future.
If you enjoyed this free chapter on the immunology, consider buying the book, for only $30.