Latest News
- Clues beginning to emerge on asymtomatic SARS-CoV-2 infection
- Back in November of 2020, during the first wave of the COVID-19 pandemic, I was teaching an in-person microbiology laboratory. One of my students had just been home to see his parents, and they all c…
- Read more
- Could there maybe be better uses of genetics and probiotics?
- Professor Meng Dong and his laboratory have created a probiotic that can metabolize alcohol quickly and maybe prevent some of the adverse effects of alcohol consumption. The scientists cloned a highl…
- Read more
- ChatGPT is not the end of essays in education
- The takeover of AI is upon us! AI can now take all our jobs, is the click-bait premise you hear from the news. While I cannot predict the future, I am dubious that AI will play such a dubious role in…
- Read more
- Fighting infections with infections
- Multi-drug-resistant bacterial infections are becoming more of an issue, with 1.2 million people dying of previously treatable bacterial infections. Scientists are frantically searching for new metho…
- Read more
- A tale of two colleges
- COVID-19 at the University of Wisconsin this fall has been pretty much a non-issue. While we are wearing masks, full in-person teaching is happening on campus. Bars, restaurants, and all other busine…
- Read more
Chapter 15 - Immunity - Introduction and Innate Immunity
15 - 1 Introduction
From a microbe's point of view, our bodies are excellent places to live and as a result we are covered with microorganisms. Most of these associations are benign or even beneficial, but a few are damaging to the animal host and they would rapidly spread throughout our tissues if it were not for the immune system. This intricate and complex collection of proteins, cells and tissues has the daunting task of keeping the tens of billions of little travelers attached to our bodies from entering areas where they ought not to be. It is nearly unbelievable that the immune system is up to the task, yet it certainly is and our lives depend on it. This chapter will describe the array of defenses that mammals have to protect themselves from infectious disease.
15 - 2 Important Introductory Concepts of the Immune System
Learning Objectives
- Describe how the immune system evolved over time.
- Define self-vs. non-self and how this is important for immunology.
- Define an antigen and what type of macromolecules the body recognizes as antigens.
- Describe T cells and B cells as the principal players in the host immune response and how they function.
Where It Came From
Almost all eukaryotic organisms have some way of dealing with the pathogenic microbes in the environment. Even unicellular eukaryotes have mechanisms to defend themselves, although these mechanisms are very simple. In these cases, the cell either keeps the microbes outside or rapidly destroys them once they enter the cell. In many cases, these unicellular eukaryotes (e.g., Paramecium and Amoeba species) actively take up bacteria into digestive vacuoles where they rapidly kill and then degrade the bacteria, releasing the residue as food. As you will see later in this chapter, some cells of the immune system (phagocytes) have inherited this capability.
As time passed and evolution proceeded, descendants of these single-cell organisms began to group together and form multicellular organisms. Once an organism becomes multicellular, the game changes a bit. Now, in many cases, there are places inside the organism that must be protected from microbes. Many small worms have a digestive tract and it is undesirable for them to have certain microbes grow there. To this end, the worms secrete a sticky substance that glues the microbes to the sides of the tract, causing them to be pushed along the tube and excreted. Somewhat distantly related systems (e.g., production of antibodies) serve a similar purpose in our bodies.
The above systems are crude and not very effective and many individuals of these species fall victim to disease. Primitive multicellular organisms assure the survival of the species by producing many progeny. As long as sufficient numbers survive and reproduce, the species continues to exist.
Further down the time-line of life on this planet, the number of cells in organisms continued to increase and so did the size and complexity of the animal. Being a large animal has advantages, such as being better able to defend yourself against predators, having the option of eating smaller folks around you and being better able to forage for food. However, it takes longer for a large organism to reach reproductive age and therefore they need to hold off the horde of pathogens for a much longer time. Guarding the body from infection was now reliant on specialized cells. These protective cells used mechanism of their ancestors, but the much slower generation times of animals demanded the immune system be more effective and sophisticated. The most complex multicellular arrangements are found in vertebrates (e.g., mammals, reptiles and birds, all of which are relatively long-lived species) and it follows that they will have the most complex immune systems. The primitive immunity of non-vertebrates is generally called innate immunity. Vertebrates have retained innate immunity, while adding the adaptive immune response.
The point of the above discussion is to illustrate why mammalian immune systems are some of the most complex and how they have evolved over time from simpler defense systems. As each new function evolved, it was added to the capabilities that were already there. In most cases the old functions remain and still serve a useful role, resulting in mammalian systems having many layers to them. This may explain why immunity is a patchwork of different systems, some of which overlap in their duties.
The Identification of Self
A central part of the job of the immune system is to differentiate self from non-self. In other words, invading pathogens must first be differentiated from the background of the host. When the immune system fails at this function, really bad things happen, including autoimmune diseases (when the immune system attacks the host) and even death. At the macroscopic level, the difference between a human and a bacterium is obvious, but at the molecular level, this is less clear. We are made of the same basic building blocks of protein, sugars, DNA and lipids. Yet the host must be able to tell its own macromolecules from those of invading pathogens.
So how does the organism figure this out? Innate immunity and adaptive immunity do it differently. Innate immunity recognizes classes of molecules that are only found in microbes, like flagellin (a flagellar protein) or lipopolysaccharide. The recognition of these molecules then triggers a response. Adaptive immunity recognizes macromolecules that are not self and then mounts a powerful response. Below we explore these ideas in more detail.
A key to the recognition of non-self by innate immunity is the presence of receptors on the surface of cells that respond to macromolecules produced by microbes. The cell must have many different receptors to combat the many different kinds of pathogens (Gram-negative and Gram-positive bacteria, fungi, and protozoa) it encounters. The structures found on these pathogens can be quite different and it is not surprising that they are recognized by different receptors produced by the immune system. In contrast to adaptive immunity, many of the ligands that innate immunity responds to are carbohydrates and lipids.
Macrophages have many receptors like this that work in concert with the complement system to bind pathogens. Receptors that recognize carbohydrates are called lectins and two examples that are found on macrophages are the mannose receptor and the glucan receptor. Macrophages also contain one kind of a very important class of molecules, the Toll-Like Receptors (TLR) which are also called patter recognition receptors or PRRs See Table 15.1.
Table 15.1. Pattern Recognition Receptors (PRR) Found in Humans
| Toll-Like Receptor | Example Ligand | Example of pathogen recognized | Cells with PRR |
| TLR1:TLR2 heterodimer | lipopolysaccharide, lipoteichoic acid, peptidoglycan | Gram-negative and Gram-positive bacteria | Monocytes, Macrophages, Neutrophils, Dedritic cells (DC), B cells, T cells, NK cells |
| TLR2:TLR6 heterodimer | Gram-negative and Gram-positive bacteria, Yeast | lipoteichoic acid, heat shock protein 70, zymosan, others | Macrophages, Monocytes, some DCs |
| TLR3 | Viral double-stranded RNA | Viruses with double stranded RNA | DCs, B cells |
| TLR4 homodimer | lipopolysaccharide, heat shock proteins, more | Gram-negative cells, hepatitis C virus | Monocytes, macrophages, B cells, some DCs, granulocytes, and T cells |
| TLR5 | Flagilin protein | Flagellated bacteria | Monocytes, macrophages, some DCs |
| TLR7 | single-stranded RNA | Unknown | Monocytes, macrophages, B cells, some DCs |
| TLR8 | single-stranded RNA | single stranded RNA viruses, i.e. polio | Monocytes, macrophages, some DCs, mast cells |
| TLR9 | Unmethylated CpG rich DNA | Bacteria | Monocytes, macrophages, some DCs |
| TLR10 complex (homodimer or heterodimer with TLR1 and 2 ) | Unknown | Unknown | Monocytes, macrophages, some DCs |
PRRs are a family of receptors, each of which binds to a different class of microbial macromolecule. PRRs are found in invertebrates as well as vertebrates indicating that this is an ancient system for detecting infection. PRRs have two domains, an extracellular domain that binds to the target molecule and a cytoplasmic signaling domain that communicates activation of the PRR to the inside of the cell. Crystal structures have revealed that the extracellular domain of the PRR has a horseshoe-like shape, where the target binds. Humans have 12 PRR genes and each PRR will respond to a certain class of molecule present on pathogens, but not found in human cells. For example, TLR4, found on macrophages, responds to the LPS of Gram-negative bacteria and other related compounds and TLR3, found on natural killer cells, recognizes double stranded RNA; a molecule only present in some viruses. Taken together the 12 PRRs recognize features that are typical of most pathogens and thus can respond to the presence of many different infections.
When a PRR on a macrophage is activated, a response cascade ensues inside that cell that causes the production of inflammatory cytokines. These cytokines are secreted into the environment and precipitate a series of responses, most importantly being the recruitment of neutrophils to the affected area. The infiltrating cells further the inflammatory response and this is described in section 15.5. This inflammatory signal by macrophages is one of the bodies best defenses against infection and marshals the rest of the immune, both innate and adaptive immunity.
Adaptive immunity has a different approach than innate immunity. It most strongly reacts to proteins, but will also react with other macromolecules, even though these structures are made by every living thing. It's solution is to create a huge population of cells (lymphocytes) that collectively are capable of responding to many macromolecules, both those that are part of our bodies (self) and those of invading pathogens (non-self). Self, in these terms, means the proteins, sugars and lipids floating around that are part of our body and are accessible to the immune system. The key step occurs during the maturation of these lymphocytes. Those cells that respond to self-macromolecules are eliminated or suppressed, while those that do not react to self are allowed to mature. In this way, the immune system learns to tolerate the host: the assumption is that any immune cell that survives this process will respond only to non-self macromolecules. The macromolecule it reacts with may be present in the outside world, potentially part of a pathogen. If that molecule enters the body, the cells that recognize it will activate and alert the immune system, eventually raising an immune response. But how do these cells recognize macromolecules? Before we tackle that question, let us first describe these non-self macromolecules in more detail. Immunologist call these macromolecules antigens.
Antigens
Antigens are defined as any type of molecule that causes an immune response in a host by interacting with antigen-specific receptors on the membrane of host lymphocytes. Proteins by far are the strongest antigens, followed by sugars, lipopolysaccharides and then lipids and DNA. This arrangement makes some sense since it is loosely based on the uniqueness of the molecule in each species. Protein structures tend to have distinct characteristics in each organism. Carbohydrates are less distinctive, and lipids and DNA are nearly universal in structure. Therefore, it is wise for the immune system to focus on proteins when trying to identify foreign macromolecules. Host T cells and/or B cells recognize these molecules as foreign and then marshal an immune response against them.
B Cells and T Cells
So how do lymphocytes recognize these antigens? By using antibodies and T cell receptors. Antibodies are a class of proteins made by plasma cells (a type of differentiated B cell) that each contain a unique variable region that recognizes just one antigen. Figure 15.1 shows the structure of an antibody. Your body is capable of making millions of different types of antibodies and together they are capable of recognizing almost any substance that is non-self. You need this tremendous variability to respond to the numerous antigens encountered in a lifetime. B cells use membrane-bound antibodies that act as receptors for foreign antigens. When membrane-bound antibody receptors on B cells recognize their antigens, they bind to them and trigger the B cells response, with the eventual synthesis of antibodies.
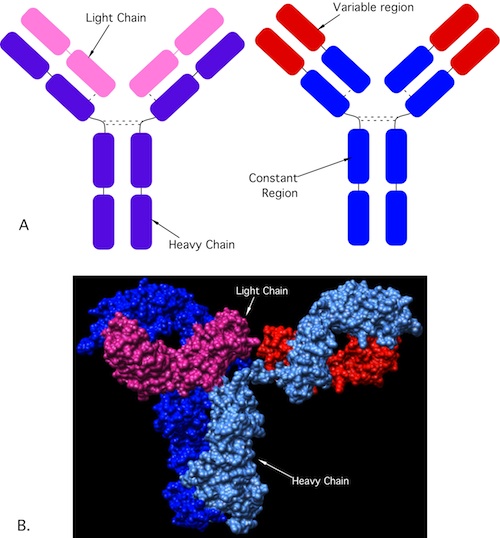
Figure 15.1. Antibody structure. (A)A cartoon of an antibody. Antibodies are composed of a light chain (pink) and a heavy chain (purple) connected by sulfhydryl linkages. The proteins combine to form a Y-shaped structure. The ends of the Y are the variable region (red) and are unique to each antibody interact with the target of the antibody, while the base (blue) is conserved and interacts with the immune system. (B) A crystal structure of an antibody showing the light and heavy chains. Source: Saphire et al. 2001. Science 293:1155-1159)
A second line of defense are T cells. These also recognize antigens through proteins on their surface called T cell receptors. T cell receptors have several antibody-like features and their binding of antigen is somewhat analogous to that seen with antibodies. Once a T cell binds an antigen, it excites the rest of the immune system to attack the source of the antigen. T cells are also the part of the immune system that destroys cancer cells and cells taken over by viral infection. The big picture is that the immune system recognizes invaders by the antigens they carry using antibodies and T cell receptors and then enlists a whole collection of cells and proteins to rid the body of the pathogen.
Key Takeaways
- Our immune system has evolved over time and involves many different systems. The patchwork nature, where many of the functions appear to overlap and address similar needs, is a reflection of this build up over time.
- The key aspect of the immune system is its ability to recognize self from non-self, and then to inactivate the non-self entity.
- The molecular representation of this non-self is the antigen. An antigen is any molecule that raises an immune response and this is most often a protein.
- Once detected, a large arsenal of systems are brought to bear on any invader and this response involves two types of cells, B cells, that differentiate into plasma cells and create antibodies against antigens, and T cells, that direct the rest of the immune system. T cells are also important in dealing with cells infected with intracellular parasites and cancer cells.
15 - 3 Susceptibility to a Pathogen Varies Between and Even within Species
Learning Objectives
- Explain how each species can be more or less resistant to specific pathogens.
- Explain how even within a single species, individuals in the population can have different susceptibilities to a pathogen.
Before we begin an examination of the immune system it is worthwhile to consider whether an animal is even susceptible to a disease. While not really part of the immune system, this can still determine the outcome of an encounter of an animal with a pathogen.
Differences in Susceptibility between Species
Whether a pathogen can cause disease in a host is dependent not only on the virulence of the pathogen, but also on the genetic background and health of the host. Some species have an innate susceptibility to a pathogen not shared with other related hosts. For example, humans are the only host for the agents of syphilis, gonorrhea, measles and poliomyelitis. In contrast, we have innate resistance to canine distemper virus and feline leukemia virus. These differences in susceptibility may be related to a number of factors. The resistant host may lack a cellular receptor required by the pathogen for attachment or penetration of the host. The temperature of the host may also preclude the growth of a potential pathogen. For example, Mycobacterium tuberculosis does not cause illness in frogs since it cannot grow well at temperatures much below 37 °C. Being cold-blooded, frogs do not normally reach these temperatures. Pathogens may also require a nutrient that is not available in a resistant host. Purine-requiring strains of Salmonella typhi cannot cause disease in rats, since rats do not make purines available for the pathogen to grow on. A final possibility is the lack of a target site for a toxin that the microbe produces. Rats injected with diphtheria toxin show no ill effects because the rats do not contain a receptor on their cell surfaces that allows the toxin to enter the cells. Since the toxin cannot enter the cells, it cannot have its toxic effect.
Differences in Susceptibility within a Species
Individuals within a species can also exhibit different susceptibility or resistance to a pathogen when compared to others. The age of an individual can have an overall effect on disease resistance, with the very young and the very old being more susceptible to infection by a wide variety of pathogens. Stress in the form of extreme exertion, shock, a change in environment, climate change, nervousness or muscle fatigue can have a negative impact on health. Each of these conditions is thought to increase the release of cortisol from the adrenal cortex, causing a suppression of the inflammatory response, thereby facilitating infection.
A chronic disease or the treatment of that disease can also weaken the body of an individual and open it up to secondary acute infections. The normal immune defenses can be impaired by serious underlying illnesses, such as AIDS, Hodgkin's disease and diabetes. Treatment of many cancers involves killing fast-growing cancer cells. This has the unwanted side effect of killing fast-growing non-cancerous cells, including the ones that make up the immune system. Patients undergoing chemotherapy and radiation therapy are therefore more susceptible to infections. The increasing numbers of transplant patients, who must take immunosuppressive drugs to prevent rejection, are also at higher risk for infection than the general population.
Poor diet is another factor that can suppress the immune system. A number of studies link vitamin and protein deficiencies to a higher rate of infection and general immune suppression. Malnutrition is in part responsible for higher rates of infection and higher infant mortality in developing nations. Being obese is also detrimental to the overall health of an individual. Morbidly obese people are more susceptible to invasive streptococcal infections. Obesity also leads to type 2 diabetes, which in turn makes people more susceptible to infectious diseases. As you can see, eating too much of the wrong kinds of foods and becoming overweight can lead to all kinds of trouble.
There are also sexual differences in the degree of susceptibility to disease. In some cases, anatomical differences cause members of one sex to be more resistant to infection than those of the opposite sex. Obviously, men cannot suffer infections of the uterus and women do not develop prostatitis. More subtly, urinary tract infections are 14 times more likely in women, because bacteria more easily travel up the short 4 cm female urethra than the longer 18 cm male urethra to reach the bladder. In addition, because of the anatomical closeness between the opening of female urethra and anus, intestinal bacteria easily gain entry into their urinary tract.
Differences in genetic background between individuals can also have an influence on susceptibility. For example, Eskimos, Native Americans and Asians are more susceptible to tuberculosis than are Caucasians. Also, individuals that are heterozygous for the gene that causes sickle-cell anemia are more resistant to the malaria protozoan. Another example comes from the AIDS epidemic. There are rare cases of individuals who have had frequent unprotected sexual encounters with virus-carrying partners yet have not contracted the disease. It turns out these individuals have a mutation in a cellular surface protein that makes it impossible for the AIDS virus to enter their cells. Without the presence of the normal surface protein, the virus cannot infect and cause disease.
Key Takeaways
- Not every pathogen is able to cause disease in every organism. In fact, pathogens can be incredibly specific, some only infecting a single species.
- On the molecular level this is dictated by the host having an appropriate gateway for the pathogen to bind to and maybe enter the host, and then also having the nutrients or molecular machinery that the pathogen requires to replicate.
- Even within a species, not every individual is susceptible to every disease. Differences in genetic background, age, sex, weight, and diet can all increase or decrease the likelihood of an illness.
- Finally, suppression of the immune system, by drug treatments or by infection with another agent, can influence susceptibility.
15 - 4 The Immune System Is a Complex Collection of Organs, Tissues, and Cells
Learning Objectives
- Define the primary tissues of the immune system.
- Define the secondary tissues of the immune system and their function.
- Explain the function of the circulatory and lymphatic system in the immune system.
- Understand the role of the bone marrow in the immune system.
- Describe the role of phagocytes, polymorphonuclear cells, mast cells, monocytes, macrophages, and lymphocytes, in the immune system.
- Describe the function and structure of the lymph nodes, MALT and the spleen.
The Immune System Is a Complex Collection of Organs, Tissues, and Cells
The immune system consists of a complex network of organs and tissues, connected by blood and lymphatic vessels, that work together to prevent infection. Many of these systems are unleashed by activation of other parts of the immune system. The extensive interdependence of the players of the immune system can make it difficult for the beginning student to understand. As we go through the immune system, you will encounter unfamiliar terms, but be patient because they will be explained later in the chapter. To try to make it a bit easier, we will first describe the anatomy of the immune system, then examine the various cell types involved in immunity, and finish by describing how these parts work together in reacting to an invading pathogen.
Many different organs and tissues in the body contribute to the function of the immune system. These include the circulatory system, bone marrow, thymus, spleen, lymphatic system and Mucosal Associated Lymphoid Tissue (MALT). Together these tissues are responsible for the creation, transport and successful operation of mammalian immunity. In this section we will look at the role of each tissue and introduce a number of cells and proteins involved in immunity.
The tissues of the immune system fall into two groups based upon their role in host defense. Primary (or central) tissues look after immune cells, creating and educating them during their differentiation into mature cells. The bone marrow and thymus gland are parts of the primary immune system. Secondary (peripheral) immune organs look after mature cells that are an active part of defense. The secondary system encompasses the rest of the immune tissues: the spleen, the lymphatic system, lymph nodes and MALT. Of course it is not this simple and the spleen and MALT also help in the maturation of immune cells. Figure 15.2 shows the anatomy of the blood and lymph systems.
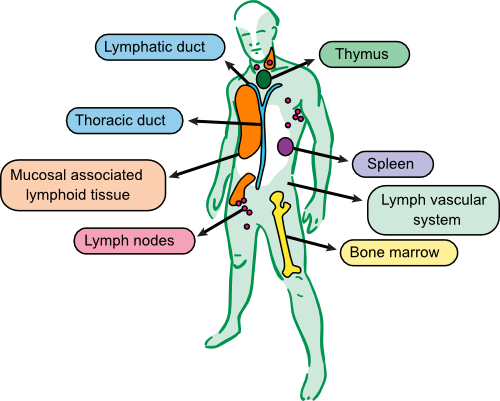
Figure 15.2. The tissues of the immune system. A large collection of organs and tissues are involved in the immune system of animals. Some of these tissues create and or educate the immune system, bone marrow and thymus, while other parts are involved in fighting infections, lymph systems, lymph nodes, spleen and MALT. The thoractic duct collects liquid from the lymph system and returns it to the circulatory system at the left subclavian vein near the heart.
The Circulatory System Transports the Components of the Immune System
The circulatory system is responsible for the transport of blood throughout the body and consists of the heart, the lungs and the vascular system of arteries, capillaries and veins. Blood, which runs through this vascular system, contains both cellular and non-cellular components. The major cell type found in blood are red blood cells, whose role is to transport oxygen and carbon dioxide into and out of the body respectively. The circulatory system also plays a secondary role as one of the routes immune cells use for transport around the body. Blood contains many types of what are called white blood cells, which are made up of mainly neutrophils, but also monocytes, T-lymphocytes and B-lymphocytes.
The non-cellular portion of the blood is a liquid called plasma. A major constituent of plasma is immunoglobulins (antibodies). It also contains various proteins of the complement system, which are equally important in the immune system. Plasma also contains the protein fibrinogen as a major component. Under the right conditions, fibrinogen participates in a complex series of reactions that eventually result in the formation of a fibrin clot in the blood. Platelets also participate in this process. Blood clots are important in stopping bleeding and also in inhibiting the invasion of advancing pathogens by entrapping them.
Bone Marrow Is the Origin of All Immune Cells
The bone marrow is the source of many immune and blood cells in the healthy adult animal. If the bone is split lengthwise, a marked difference in tissue is noticed. Part of the tissue is red, which is the source of red and white blood cells. The other portion is yellow adipose tissue that is inactive. During an infection, the yellow marrow can be reactivated to become red marrow to help in the production of larger numbers of immune cells.
In the adult animal, all immune cells originate from hematopoietic stem cells located in the bone marrow. Stems cells constantly divide and differentiate into various types of immune cells under the influence of cytokines (cytokines are small signaling proteins that help to regulate the behavior of the cells of the body). Figure 15.3 shows the origin of the cells of the immune system. The bone marrow is ultimately responsible for the synthesis of eight types of cells: red blood cells, platelets (through megakaryocytes), neutrophils, basophils, eosinophils, mast cells, monocytes/macrophages, T lymphocytes (T cells) and B lymphocytes. Some of these cell types mature in the bone marrow itself, while others migrate through the circulatory system and undergo final maturation in other tissues. At this point, we will take a short stop and look briefly at some of the features of the cells created in the bone marrow as an introduction to their function. You will learn more details about each immune cell type later when we cover specific immune responses.
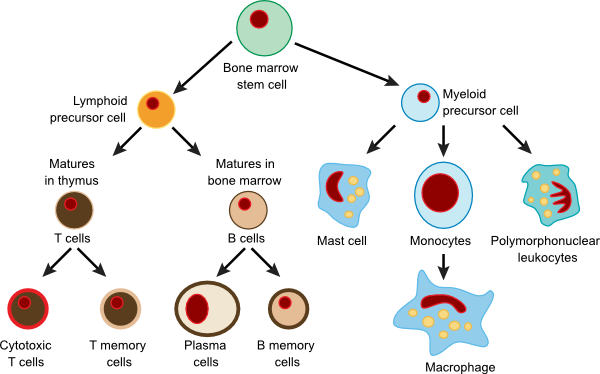
Figure 15.3. Types of immune cells and their progenitors. In humans stem cells differentiate into different types of immune cellsin the bone marrow under the influence of different cytokines.
Cells Made in the Bone Marrow
Hematopoietic stems cells create a large number of red blood cells. The heathy human creates 2.4 million red blood cells per second and they are essential for moving oxygen in and carbon dioxide out of our system. Another cell type produced in the bone marrow are megakaryocytes. These then differentiate into platelets that are necessary for the formation of blood clots following any kind of injury. Blood clots are vital in stopping the flow of blood when blood vessels are injured, but also have a role in fighting infections (see below).
Polymorphonuclear granulocytes is the general term given to neutrophils, eosinophils and basophils. The first half of the name describes the appearance of the nucleus that seems to be split into a number of different lobes. In reality, the nucleus is contiguous, but contains many infoldings, which give it a polynuclear appearance. The rest of the name comes from the appearance of the cytoplasm, which looks speckled. The cytoplasm is full of granules that contain compounds and enzymes important in fulfilling the function of each cell type. Polymorphonuclear granulocytes make up 50-70% of the white blood cells found in blood. They last only about three days and have to be replaced at a rate of 80 million cells per minute.
Neutrophils are the most common type of polymorphonuclear cells, making up 90% of granulocytes in the blood. These cells function as phagocytes in attacking and destroying infectious agents. We will cover their roles in more detail when we discuss phagocytes.
Eosinophils make up 2-5% of granulocytes in the blood, but this number can rise considerably in people with parasitic diseases as well as asthma, eczema or other diseases associated with allergies. They are primarily found in the blood, but also near epithelia that have high bacterial populations (e.g., intestines, vagina, nasal passages). The granules in these cells bind the red dye eosin, giving the cells their name. Eosinophil granules contain a number of different enzymes including, acid phosphatase, glucuronidase, cathepsins, RNase, and arylsulfatase and peroxidase. They also produce toxic basic proteins. They respond to the chemical signals put out by other immune cells and can then participate in an immune response. The major reactions take three forms.
- They can down-regulate an immune response by destroying histamine secreted by mast cells using the enzyme histinase. Eosinophils also liberate arylsulphatase that breaks down the slow reactive substance of anaphylaxis (a dangerous form of allergic response) that is released by mast cells.
- Eosinophils combat antigenic challenges too big to be attacked by phagocytes. Examples of such challenges are parasitic worms or helminths. In battling these infections, the body first covers the worm with antibody. This then activates eosinophils, which bind to the parasite and release the contents of their granules, thus causing external digestion of the worm.
- As is the case with neutrophils, eosinophils can phagocytize microorganisms, but this is a secondary role.
Basophils are small cells that make up less than 1% of all white blood cells. The granules of these cells contain heparin, histamine, decarboxylase, histidine, dehydrogenase and diaphorase. Heparin is an important anti-clotting compound, and histamine finds its use modulating the immune response. Histidine is converted to histamine by decarboxylase. The role of basophils in the immune response is not yet clear, but they seem to play a role in the defense against parasitic worms and in severe allergic reactions. They have a very high affinity for one class of antibodies (IgE) and they are usually found coated with IgE in tissues. Binding of IgE may set in motion a series of events that causes other immune cells to respond to the high concentrations of IgE. Basophils may be cellular alarms that notify the rest of the immune system and help to concentrate the point of attack.
Mast cells are closely related to basophils but are distinct in their reactions to antigens. They are found throughout the body in lymph nodes, spleen, bone marrow, around blood vessels, nerves, glands and in the skin. Mast cells have granules that, like basophils, contain heparin and histamine. They have a high affinity for IgE as well and their activation by antigen triggers histamine release. Until recently, they were mostly thought to trigger unwanted allergic reactions, but it is now becoming clear they participate in immune responses to Gram-negative bacteria and to nematodes, such as the hookworm. Their wide distribution indicates that they are important in many immune responses.
Monocytes and macrophages are long-lived specialized phagocytic cells. Monocytes are migrating phagocytic cells found in the bloodstream and when they enter other tissues, they differentiate into macrophages. Macrophages are found in the brain, lungs, liver, spleen, lymph nodes, joints and peritoneum. The key functions of monocytes and macrophages are to remove our own dead cells when they reach the end of their useful life and also to remove pathogens. For example macrophages in the liver, called Kupffer cells, phagocytize old erythrocytes from the blood and remove them. Another one of their functions is the creation of important immune proteins and peptides. They are responsible for synthesizing transferrin (an iron-binding protein), and various cytokines necessary for immune function. Macrophages also synthesize some of the complement proteins.
B lymphocytes or B cells can differentiate into plasma that are responsible for antibody production. They are very important in fighting many different types of infections, especially, bacterial infections. T lymphocytes are involved in regulating the immune system and destroying host cells that are out of control, either due to a breakdown in cell division regulation (cancer) or infection by a virus or even an intracellular parasite. We will discuss the functions of these cells in more detail when we cover the adaptive host response.
The thymus is a fist-sized organ located above the heart that is involved in the maturation of T lymphocytes (also referred to as T cells). T cells produced by the bone marrow are immature and journey to the thymus through the bloodstream. The blood vessels that supply the thymus with oxygen and other nutrients also contain a blood-thymus barrier that only allows immature T cells in and mature T cells out. The thymus is also connected to the lymphatic system through lymph vessels. We will talk more about thymus function when examining T cell maturation later in the chapter.
The Lymphatic System Is a Separate Vascular System Through Which Lymph and Immune Cells Move
The lymphatic system is a separate vascular system, distinct from the bloodstream, through which the lymph moves. It is a branching system whose vessels get ever smaller as it penetrates tissue. Unlike the blood system, the lymphatic system is not circular, instead its capillaries extend into tissues and eventually having closed ends. The system focuses its attention on areas of the body that are most likely to be entry points for pathogens: skin dermis, respiratory tract, gastrointestinal tract and genitourinary tract. It functions are:
- To collect excess fluid from surrounding tissue and return it to the bloodstream
- To absorb fat from the villi of the small intestine.
- For the transport of Immune cells moving through the body
Here we will focus on its major roles of the lymph system, harboring and transporting many of the cells involved in the immune system. Lymph fluid consists of leukocytes and many components of plasma, but does not contain red blood cells. Liquid enters the lymph system from non-vascular tissue draining into lymph capillaries. The capillaries then drain into lymph nodes that sit at the junction of a number of lymph vessels.
Lymph nodes, shown in Figure 15.4, are found throughout the body and are major centers for immune function. In a healthy adult the smallest lymph nodes are a few millimeters in diameter, while the largest can reach 2 cm. Large volumes of liquid and cells pass through lymph nodes each day and are filtered to detect antigens and remove microbes. The nodes also interact with phagocytes to begin various immune functions that we will elucidate later. Lymph nodes are dynamic, densely packed structures with the bulk of the cells inside them being mobile. Phagocytes and antigens enter through as many as five vessels, called afferent ducts. The lymph node contains large numbers of B- and T-lymphocytes, but macrophages and plasma cells are also present. Macrophages carry out the job of filtering the incoming fluid and plasma cells (effector cells that differentiate from antigen-stimulated B cells) secrete antibodies that exit along with other immune cells by the single exit, the efferent duct. The lymphatic system then merges into larger vessels and eventually reconnects to the circulatory system through ducts near the heart. The flow of liquid is therefore from the lymphoid capillaries, into lymph vessels and eventually exiting into the bloodstream at the heart. There are actually two asymmetric lymphatic drainage systems, the both drain into veins near the heart. The right lymphatic duct drains the upper right side of the body, including the right side of the head, the heart and lungs, into the right subclavian vein. The rest of the body drains into the thoracic duct, which then drains into the left subclavian vein. Liquid entering the lymphatic system from the blood and extravascular tissue filters through typically eight to ten nodes before returning to the bloodstream.
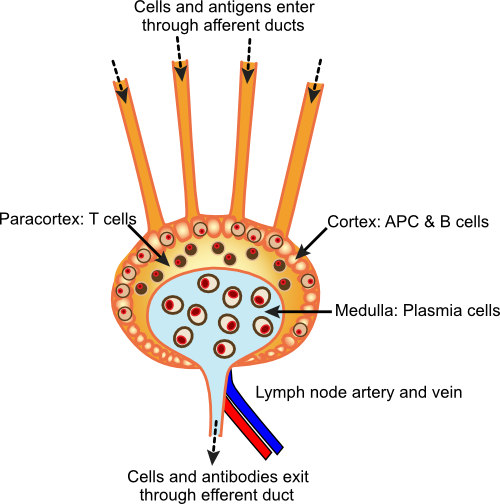
Figure 15.4. A diagram of a lymph node. Lymph nodes contain many types of immune cells organized to monitor and react to antigens that come in from the lymph system. T cells and B cells line the edges of the lymph node and plasma cells and phagocytes are found in the core.
The MALT and Spleen Are Collections of Immune Cells That Work to Fight Infection
Mucosal-Associated Lymphoid Tissue (MALT) is scattered throughout the connective tissues of the body, but especially beneath moist epithelial membranes such as those that line the upper respiratory tract, intestine and urinary tract. MALT is strategically distributed to help the body prevent infection by organisms that have penetrated beyond the mucosal surface. MALT consists of small masses of lymphatic tissue (up to a millimeter in diameter) containing mainly lymphocytes. These tissues are far less organized than the lymph nodes. Most MALT consists of small groups of cells, but in certain areas it is found in large clusters. For example, large aggregates of MALT occur in the wall of the lower portion (ileum) of the small intestine and are known as Peyer's patches. Tonsils and adenoids are also aggregates of MALT that protect your body from microorganisms present in the upper respiratory tract.
The spleen is a very important secondary lymphoid organ. Individuals who have had their spleens removed due to rupture caused by a car accident or atrophy from sickle cell anemia can lead nearly normal lives, but they tend to be more susceptible to infection. Some functions of the spleen are similar to those of the lymph nodes; however, it also produces lymphocytes and removes senescent (old) red blood cells from the circulation.
The spleen, shown in Figure 15.5, is highly organized and is a repository for immune cells. It plays an important role in response to pathogens. The spleen contains a circulatory system allowing access to it through the blood. Cells in the spleen organize around the blood vessels into two tissues: red pulp and white pulp. Red pulp contains mainly red blood cells, and white pulp is made up predominantly of lymphocytes. The white pulp focuses tightly around the arterioles of the spleen while the red pulp fills the rest of the interstitial space. The location of various lymphocytes in the white pulp further differentiates it. T cells are found near the arterioles and further away are areas of B cells. The spleen is a major area for B cells to congregate in the body, where they wait to be activated by antigens. During an illness, activated lymphocytes are released from the spleen to fight the infection.
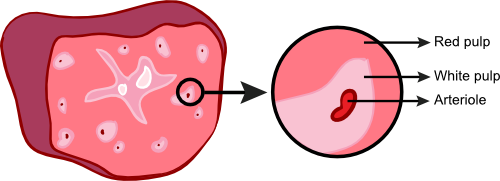
Figure 15.5. Anatomy of the spleen. The spleen is located on the middle of the abdomen to the left of the stomach. This cartoon of a cross section of the spleen shows both red and white pulp. The magnified section shows the arrangement of cells. The white pulp near the arterioles consists of T cells and B cells and the red pulp is filled with red blood cells.
This completes our tour of the anatomy of the immune system. In the next section we will more specifically examine how cells and compounds in these tissues recognize and attack invading pathogens.
Key Takeaways
- Many different tissues distributed throughout the body are involved in protecting us against microorganisms. The immune system can be divided into primary (or central) tissues: the bone marrow and thymus, and secondary (or peripheral) tissues; the spleen, the lymphatic system, lymph nodes and MALT.
- Primary tissues create and educate immune cells, while secondary tissues monitor the body for invading pathogens.
- The bone marrow creates phagocytes (neutrophils, basophils, monocytes and macrophages), mast cells and lymphocytes, all of which are important in the immune response.
- Phagocytes ingest and kill cells identified for destruction by the immune system.
- Mast cells are present in many immune tissues and react to incoming antigens, alerting the rest of the immune system.
- Lymphocytes are divided into two forms, T cell and B cells.
- Lymph nodes and the lymph system are a secondary vascular system that monitors the body for antigens. The lymph node contains T cells, B cells and macrophages.
- The spleen is also a reservoir of B cells, T cells that are released when activated to fight an infection.
15 - 5 The Innate Immune System
Learning Objectives
- Define the innate immune system and its role in protection against pathogens.
- Be able to list the structures, secretions, and microorganisms (yes microorganisms) involved in innate immunity.
- Innate immunity differs from adaptive immunity in that
Host defenses can be divided into two categories, innate and adaptive immunity. Innate immunity involves general mechanisms in a healthy animal that prevent colonization by microorganisms and antagonize or kill those that do enter the host. They are always present and the strength of their response does not increase with repeated exposure to the inducing microbe. Adaptive immunity develops through the mechanisms that are turned on in response to a pathogen. This involves activation of the immune system, where the initial response to a pathogen is weak but becomes quite vigorous over a period of a few days. Adaptive defenses also have a memory of encountered pathogens such that a second infection by a pathogen is met with a more rapid and vigorous immune response. The rest of this chapter will cover innate immunity. The second chapter will describe adaptive immunity, summarize with a description of the entire immune systems response to a pathogen, and then finish by presenting errors the immune system makes and how those affect the body.
Innate immunity is the first line of defense against a pathogen. The system must be somehow circumvented by the pathogen before it can enter into the host. In most cases, the adaptive immune system is only activated after the innate immune system has been breached. Here we examine various structures, products and cells of the innate response as shown in Table 15.2.
Table 15.2.The Innate Immune System
| Defense | Mode of Antagonism |
| Anatomical structures (skin, hair, blinking, nose hairs, cilia in the lungs and the peristaltic action of the digestive track) | Provide physical barriers or motion that sweeps microbes out of areas of the body |
| Tissue bactericides | Many proteins and chemicals are created by the body that kill or inhibit the growth of microbes |
| Microbial antagonism | The normal flora of the body prevent pathogens from colonizing and causing disease. |
| Complement proteins | These proteins can be triggered by microbial secretions or by antibodies. They alert the immune system and can cause cell lysis. |
| Inflammation | A reaction to tissue damage that involves a large collection of cells, proteins and chemicals. |
| Phagocytes | Cells that attack microorganisms, engulf them and kill them. They are a major defense of the body. |
| Natural killer cells | Cells that roam the body and attack cells coated in IgG. |
Parts of the immune system that are always present and whose reaction against a pathogen does not increase with exposure to it. Natural killer cell activation does depend on antibodies, but they are not inducible. Therefore we include them here.
Microbial Resistance in Anatomical Structures
Figure 15.6 summarizes the various anatomical defenses of the body. The skin is an extremely effective barrier to microorganisms. Besides preventing the cells of our body from escaping, it also prevents microorganisms from getting in. Skin contains several layers of tightly packed, heavily keratinized cells (keratin is a fibrous protein that gives skin, hair and nails its toughness). It is very difficult for most organisms to squeeze in between skin cells. Skin cells are also continually being shed and replaced by new ones, thus removing any microorganisms attached to them. The skin surface is also very hydrophobic and dry, which prevents the growth of many microorganisms. Sebaceous glands are present throughout the skin and they secrete hydrophobic oils that further repel water and microorganisms. The oil also helps keep the skin supple and flexible, preventing cracking that might allow microbial access to internal layers. Finally, melanin in the skin also helps to reduce the harmful impact of UV light by absorbing it. UV light can be damaging to all cells, including cells of the immune system.
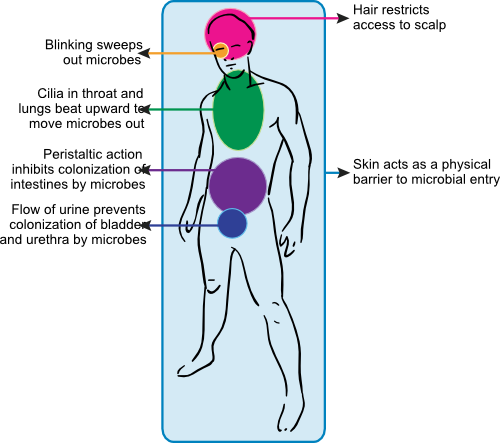
Figure 15.6. Anatomical immunity. Parts of the body are designed to prevent the passage of unwanted microorganisms. Some examples of structures that inhibit the advance of pathogens include the skin, hair, blinking eyelids, nose hairs, cilia in the lungs and the peristaltic action of the digestive track.
Hair helps to restrict access of airborne pathogens to the skin. It protects the most sensitive or exposed body orifices, including the nasal cavity, the eyes and ears. Hair also serves as a cushion lessening the severity of cuts and grazes, which decreases the depth and number of breaks in the skin.
The movement of various body parts can help to rid the body of microorganisms trying to colonize. Blinking the eyelids constantly sweeps microorganisms out of the eye. The entire respiratory tract, except bronchioles and alveoli, contains a mucociliary escalator consisting of mucus producing goblet cells and ciliated epithelium. The cilia continually beat upward and any particles that get stuck in the mucus are pushed out of the lungss. The peristaltic action of the gut not only moves food along our digestive tract, but also flushes microbes out of our system. If microorganisms do not have a method of attachment, they rapidly leave the gastrointestinal tract. The constant flushing of the urethra by urine helps to keep the bladder free of bacteria. All of the above movements continually remove microbes from our bodies.
Antimicrobial Secretions
The body also produces a number of antimicrobial substances that inhibit or kill microorganisms. Collectively these substances are known as tissue bactericides. Tear ducts, sebaceous glands, ears, nose, lungs, mouth and digestive tract all secrete antimicrobial substances. Table 15.3 lists some of the common tissue bactericides, their mode of action and location.
Table 15.3. Common Tissue Bacteriocides
| Substance | Common sources | Chemical composition | Activity |
| Lysozyme | Serum, saliva, sweat, tears | Protein | Bacterial cell lysis |
| Basic proteins and polypeptides (histones, β-lysins and other cationic proteins, tissue polypeptides) | Serum or organized tissues | Proteins or basic peptides | Disruption of bacterial plasma membrane |
| Lactoferrin and transferrin | Body secretions, serum, organized tissue spaces | Glycoprotein | Inhibit microbial growth by binding (withholding) iron |
| Peroxidase | Saliva, tissues, cells (neutrophils) | Protein | Act with peroxide to cause lethal oxidations in cells |
| Fibronectin | Serum and mucosal surfaces | Glycoprotein | Bind to bacteria and assist in clearance (opsonization) by phagocytes |
The human body produces a large number of antimicrobial chemicals and proteins to keep microorganisms in check.
One important group of broad-spectrum antimicrobial peptides is the defensins, which participate in the host defense of mammals, birds, plants and insects. Their presence in a wide variety of species probably indicates that they are an ancient form of antimicrobial antagonism. Defensins form a family of cysteine-rich, cationic and structured polypeptides of 29-42 amino acids that contain three or four disulfide bridges. They work by disrupting the membrane of a wide variety of pathogens, including Gram-negative and Gram-positive bacteria, fungi and some enveloped viruses. They also serve as chemokines, which attract elements of the adaptive immune system. Defensins are expressed by many different tissues and it is now clear that they are a vital part of innate immunity.
Microbial Antagonism
- As mentioned in Chapter 14, the normal non-pathogenic microorganisms that live inside and on our bodies as part of normal flora have an important role in defending us against pathogens. This is accomplished in three ways.
- The normal flora are well adapted to the tissues they live on and they out-compete pathogenic microbes for these sites of colonization, thus preventing harmful bacteria from gaining an initial foothold on our bodies.
- The normal flora produce specific antimicrobial compounds (called bacteriocins) that kill or inhibit closely related species. The normal flora also produce non-specific compounds such as the end products of their metabolism that may inhibit pathogenic bacteria.
- They utilize nutrients, limiting their availability to pathogens.
Complement Is a Set of Proteins That Detect and Kill Microbes
Complement is an enzymatic system in the blood containing over 20 proteins (some of the more prevalents proteins have been given the names C1-C9) . These proteins are produced in a variety of cells, including macrophages, hepatocytes (located in the liver) and epithelial cells in the gastrointestinal mucosa. They then circulate in the bloodstream until activated. Figure 15.7 shows a conceptualization of the complement cascade.
Figure 15.7. The complement cascade. The reactions of the proteins of the complement cascade. See text for details.
Activation of the complement system can follow one of three pathways: the classic pathway, the lectin pathway and the alternative pathway. The classic pathway begins when antibody of type IgG or IgM reacts with antigen (antibody structure is described below). IgG and IgM contain a masked C1 binding site that is exposed upon binding to antigen. C1q binds to this site and begins the complement cascade as shown in Animation 15.1. Binding of C1q activates C1r that in turn activates C1s. This forms an activated C1qrs enzyme that catalyzes the cleavage of C4 into C4a and C4b. Active C1qrs also degrades C2 into C2a and C2b. This is an enzymatic reaction, in which the binding of C1 to antibody results in the digestion of a large number C2 and C4 proteins. C2b and C4a are released into the environment. C2a and C4b bind to the membrane and associate to make a C3 convertase that acts on a large number of C3 proteins making C3a and C3b. C3b then combines with C4b and C2a to form a C5 convertase, cleaving C5 to C5a and C5b. The end result is the active degradation of a large number of complements proteins, by this expanding cascade. In this way a single antibody-antigen reaction can be amplified and produce an aggressive host response. Complement activation leads to several ultimate results.
- The C3a and C5a produced serve as a chemoattractants for phagocytes.
- C3a and C5a also bind to specific receptors on mast cells inducing degranulation and the release of inflammatory substances.
- C3b remains bound to the Fc portion of the antibody or may bind to the cellular membrane of the trigger. In either case, it serves as a protein that phagocytes can bind to and then envelop and kill the bacteria.
- Several of the complement proteins (e.g., C5b, C6, C7, C8 and C9) form a pore in the membrane of the initiating microbe that causes leakage of cell contents and results in cell death.
- When inserted in membranes, C8 and C9 form a phospholipase that can destroy membranes of the pathogen. This can inactivate a virus and remove the outer membrane of Gram-negative bacteria, making their peptidoglycan accessible to lysozyme.
The lectin pathway is very similar to the classical pathway except the trigger is different. The host will produce a protein called mannose-binding lectin (MBL) that can attach to polysaccharides that contain the sugar mannose. It turns out that a significant number of microbes produce mannose containing polysaccharides on their surface, and MBL will associate with them. When it does, two other proteins (MBL-associated serine protease 1: MASP-1) and MASP-2 binds to MBL forming a structure similar to activated C1qrs and precipitate the complement cascade by cleaving C2 and C4. From here the reactions are similar to the classic pathway.
The alternative pathway of complement activation does not involve antigen binding to antibody. This pathway involves interaction of the C3 compliment protein with the cell surface of a pathogen. C3 in the blood spontaneously hydrolyzes a thioester bond at a slow rate to form C3(H2O). This change in shape allows the binding of plasma protein Factor B, which is then a target for Factor D to cleave Factor B into Ba and Bb. Bb remains part of the C3(H2O) complex, now called C3(H2O)Bb. This complex, also known as a fluid-phase C3 convertase, will cleave a C3 molecule and cause C3b to interact with a membrane surface. If the membrane surface is a host membrane, it contains proteins that will short circuit the cascade. The C3a and C3b proteins rejoin, ending the reaction. If bound to a pathogen, they lack the proteins to stop the reaction and further activation of this membrane bound C3b molecule by Factor B and Factor D results in the formation of a membrane-bound C3 convertase. This convertase with activated Factor B and D is unstable, but can be become stable by the binding of properdin. Properidin is another blood protein. Once active and stable, C3 convertase digests C5, forming C5a and C5b. Subsequent steps in the alternative pathway are similar to the classical pathway. The alternate pathway allows antibody-independent activation of the complement system, which is important in initial defenses against a new pathogen before antibodies have been synthesized. Individuals who are deficient in complement formation have compromised immune systems. For example, complement deficiency in C3, C5, C6, C7, C8 or C9 leaves the person more susceptible to dangerous bacterial infections.
Key Takeaways
- The innate immune system is always present and cannot be induced by exposure to pathogens.
- Innate immunity consists of anatomical structures, antimicrobial secretions, the normal flora, complement, the inflammation response and phagocytes.
- Complement is activated by antibodies or certain microbial secretion and causes a cascade of enzymatic reactions.
- The results of these reactions is induction of inflammation, recruitment of phagocytes and the formation of a pore in the membrane of the microbial cell.
15 - 6 Inflammation and the systems that react to it
Learning Objectives
- Be able to described the events of inflammation.
- Explain the triggers of inflammation.
- Explain the role of phagocytes in inflammation.
- Describe in detail the events of phagocytosis and the mechanisms that these cells use to detect, attach to, and kill target pathogens.
The next time you burn or cut yourself, make a note of the symptoms you observe in the damaged tissue. You will observe swelling, redness, heat and pain. Swelling is caused by the creation of gaps between the capillary cells, allowing the movement of fluid and immune cells to the damaged area. An increase in blood flow to the area causes the characteristic redness. Heat is caused by the accumulation of blood and the release of fever-inducing molecules (called pyrogens). Pain is felt in response to tissue damage and the irritation of sensory nerves in the affected area. This series of events is collectively called inflammation.
Figure 15.8. Inflammation. The inflammatory response focuses the attention of the body's repair and immune systems on an area of damage. The body attempts to repel any invading pathogens and then begins the process of repairing the damage. See text for details.
Of all the host defenses, inflammation is arguably the most important. It is one of the initial responses to the presence of an invader and focuses the immune system at the site of infection. Inflammation draws phagocytes, which attack and kill the invading microbe and interact with the adaptive immune system to create a long-term response. However, inflammation can cause considerable damage to host tissues, which can be part of the microbial pathogenesis of a disease process. In addition, unwanted inflammation can also be a major destructive force, as seen in many autoimmune diseases.
Inflammation can be triggered by damage to host tissue, by certain immunological reactions (such as the complement cascade discussed previously) or by the presence of harmful microbial as well as non-microbial agents. No matter what the cause, induction and maintenance of inflammation follows a similar series of events as shown in Figure 15.8.
- Injured or dead cells release their cytoplasmic constituents in the surrounding area causing a lowering of the pH.
- The drop in pH leads to the activation of the extracellular enzyme kallikrein that in turn activates bradykinin.
- Bradykinin binds to receptors in the cells of nearby capillary walls, opening up intracellular spaces that allow blood constituents, including immune cells, access to the area.
- Bradykinin also binds to mast cells that are found in association with the small vessels of most tissues. This binding precipitates the release of the mediators of inflammation such as histamine, heparin, prostaglandins, leukotriene and other compounds.
- This cocktail of chemicals recruits neutrophils, macrophages and leukocytes that destroy the invading particles and help to induce a specific immune response.
One of the major results of inflammation is the isolation of the pathogen in a defined area. This often occurs through the formation of a blood clot - the same kind of clot that forms when the skin is cut. This response is a rapid and extremely powerful antimicrobial activity that successfully deals with many infections.
Phagocytes Engulf and Kill Foreign Cells
An important result of inflammation is the recruitment of phagocytes. These cells function to engulf and attack particles in the host that have been signaled for removal by various mechanisms. In this section we describe the synthesis and properties of phagocytes.
Phagocytes originate in the bone marrow from stem cells that first differentiate into myeloid precursor cells. These are a set of intermediate cells that can take a number of paths. Under the influence of various cytokines, the myeloid precursor cells then differentiate into different kinds of phagocytic cells such as polymorphonuclear granulocytes, monocytes and macrophages as shown in Figure 15.9. Polymorphonuclear granulocytes (mostly neutrophils) leave the bone marrow and circulate in the bloodstream and body for only a few days before dying. They are the first phagocytic cells that normally encounter an infection and can rise to large numbers during a severe illness. Monocytes differ from polymorphonuclear granulocytes in that they have an unsegmented nucleus and are much longer lived. This longer existence is critical to their function, as you will see later. After creation in the bone marrow, they circulate in the blood for a period before settling in a tissue and maturing into macrophages. Finally, macrophages are mature monocytes that are attached to lymph tissues and can be up to ten times the size of monocytes. All phagocytes contain membrane vesicles filled with destructive and degradative compounds (lysosomes) that are available for the annihilation of engulfed microbes.

Figure 15.9. Phagocytes, Neutrophils, Macrophages and Monocytes. Phagocytes are the cellular sentries of the immune system, detecting, engulfing and killing pathogens in our bodies. They also destroy dead or dying cells and cancerous cells.
Phagocytosis
Phagocytes are motile and roam throughout the bloodstream, the lymphatic system and non-vascular tissue in search of particles to assault. When a non-self particle is encountered, it is taken into the phagocyte, combined with destructive compounds and destroyed. This process, termed phagocytosis, involves several discrete steps as listed below and diagrammed in Figure 15.10.
- Detection of the foreign particle and movement of the phagocyte to the area.
- Attachment of the foreign particle to the phagocyte.
- Engulfment or ingestion of the foreign particle into a vesicle called a phagosome.
- Fusion with lysosome and formation of the phagolysosome.
- Intracellular killing and digestion.
- In the case of macrophages, egestion and antigen presentation.
Figure 15.10. The process of phagocytosis. The steps of phagocytosis, detection/chemotaxis, attachment, engulfment, fusion, and killing. Macrophage are also capable of egestion and antigen presentation.
For the sake of simplicity, we will focus on the example of the attack of a phagocyte on a microorganism. However, the process is similar when a phagocyte attacks a virus or other foreign particle.
Phagocytes Can Detect Pathogens and Move Toward Them
The presence of an infecting microbe sets into motion a number of host responses, one of them being inflammation. This in turn regulates adhesion of phagocytes to blood vessel walls that allow the migration of phagocytes across vascular walls. Once inside an infected tissue, phagocytes must then detect the location of a microbe, which can be signaled in a number of ways.
- Many microbial proteins contain N-formyl methionine at their amino terminal end, while eukaryotic proteins do not. Certain N-formylated bacterial peptides serve as chemoattractants for phagocytes.
- The binding of antibodies to antigens triggers the classic complement pathway with subsequent release of C3a and C5a. These proteins are also chemoattractants for phagocytes.
- The complement cascade also induces an inflammatory response, whose mediators of inflammation also draw phagocytes to the area.
From these three examples, it is obvious that the body creates a number of strong signals in response to microbial invasion, and that these signals quickly draw phagocytes to the area of infection. The rapid removal of microbes is critical in preventing a disease and having multiple attractants insures a swift reaction.
Attachment
The phagocyte must then attach to the target microorganism. Without assistance, this can be a somewhat inefficient task for the phagocyte. In the absence of help, phagocytes are still capable of binding to microorganisms by a mechanism that can be thought of as nonspecific attachment. This nonspecific attachment probably involves electrostatic or hydrophobic attraction between the phagocyte and microorganism. It is also possible for the phagocyte to physically trap the microbe against a tissue surface and initiate ingestion—a mechanism called surface phagocytosis. These nonspecific mechanisms of attachment are probably important early in infection to slow the microbe's progress, but they will not stop an infection.The immune system has a number of ways that make the process of phagocytosis more efficient. Phagocytes have over 40 specific types of receptors on their cell surfaces. Some of these receptors are for chemoattractants that draw phagocytes to an area; others recognize molecules that enhance the binding of phagocytes to their targets. This enhancement of binding is termed opsonization and macromolecules that bind to a microbe and increase the efficiency of phagocytosis are opsonins. Opsonins provide molecular handles for the phagocyte to grab onto. Several different macromolecules can serve as opsonins for phagocytes. IgG antibodies have a site on their constant region that can react with an antibody binding receptor on phagocytes. This phagocyte-binding region is masked in a free antibody, but becomes accessible to phagocytes when the antibody binds to antigen. Initially, masking this site prevents free antibody from binding to phagocytes. The complement protein C3b can also serve as an opsonin. Whether triggered by the classic, lectin or alternative pathway, C3b eventually binds to the membrane surface of the complement-activating microbe. Phagocytes contain a C3b receptor, which binds to C3b and thus to the microbe containing it. Opsonization of bacteria greatly increases the rate of attachment and ingestion by phagocytes. Bacteria in the blood are quickly cleared only if they are bound by opsonins.
Ingestion
Attachment of the microbe to the phagocyte results in some sort of signal (the nature of which is still not clearly understood) that triggers ingestion of the microbe. Ingestion involves encircling the target particle with phagocytic membrane so that it is eventually taken inside the cytoplasm of the phagocyte engulfed in a membrane vesicle called a phagosome. This process requires ATP and is triggered by the attachment of the target to the phagocyte's cytoplasmic membrane. Contact between a microbe and a phagocyte also changes the phagocyte's metabolism from aerobic respiratory to anaerobic fermentative, with lactic acid being the final end product. The increase in lactic acid in the phagocyte lowers the pH of the cytoplasm, including the phagolysosome and this enhances the activity of many of the degradative enzymes present.
Phagolysosome Formation
The phagosome containing the microorganism migrates into the cytoplasm and soon collides with a series of lysosomes forming a phagolysosome. When the membranes of the phagosome and lysosome meet, the contents of the lysosome explosively discharge, releasing a large number of toxic macromolecules and other compounds into the phagosome. The killing processes inside the phagolysosome are confined to the organelle of the phagolysosome, thus protecting the cytoplasm of the phagocyte from these toxic activities.
Intracellular Killing and Digestion
Several minutes after phagolysosome formation, the first detectable effect on the microorganism is the loss of the ability to reproduce. Inhibition of macromolecular synthesis occurs sometime later and many pathogenic and non-pathogenic bacteria are dead 10 to 30 minutes after ingestion. The mechanisms phagocytes use to carry out this killing are diverse and complex, consisting of both metabolic products and lysosomal constituents. Each type of phagocyte (neutrophils, monocytes or macrophages) has a slightly different mix of killing methods. The killing mechanisms that phagocytes use can be organized into two broad groups: oxygen-dependent and oxygen-independent mechanisms. Figure 15.8 shows the killing mechanisms of phagocytes.
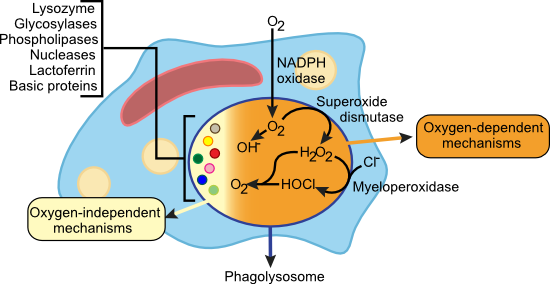
Figure 15.11. Killing Mechanisms of Phagocytes. The figure shows the many different techniques used by phagocytes to kill pathogens. Some require oxygen while others do not.
Oxygen-Dependent Mechanisms
As will be discussed later, antibodies have constant regions on them that phagocytes can bind using Fc receptors. Binding of Fc receptors on neutrophils, monocytes and macrophages (also binding of mannose receptors on macrophages) causes an increase in oxygen uptake by the phagocyte called the respiratory burst. This influx of oxygen is used in a variety of mechanisms to cause damage to microbes inside the phagolysosome, but the common theme is the creation of highly reactive small molecules that damage the biomolecules of the pathogen. Binding of these receptors activates an NADPH oxidase that reduces O2 to O2- (superoxide). Superoxide can further decay to hydroxide radical (OH.) or be converted into hydrogen peroxide (H2 O2) by the enzyme superoxide dismutase. In neutrophils, these oxygen species can act in concert with the enzyme myeloperoxidase to form hypochlorous acid (HOCl) from H2 O2 and chloride ion (Cl-). HOCl then reacts with a second molecule of H2 O2 to form singlet oxygen (1 O2), another reactive oxygen species. Macrophages in some mammalian species catalyze the production of nitric oxide (NO) by the enzyme nitric oxide synthase. NO is toxic to bacteria and directly inhibits viral replication. It may also combine with other oxygen species to form highly reactive peroxynitrate radicals. All of these toxic oxygen species are potent oxidizers and attack many targets in the pathogen. At high enough levels, reactive oxygen species overwhelm the protective mechanisms of the microbes, leading to their death.
Oxygen-Independent Mechanisms
The pH of the phagolysosome can be as low as 4.0 and this alone can inhibit the growth of many types of microorganisms. This low pH also enhances the activity of lysozyme, glycosylases, phospholipases and nucleases present in the phagolysosome that degrade various parts of the microbe. A variety of extremely basic proteins present in lysosomal granules strongly inhibit bacteria, yeast and even some viruses. In fact, a few molecules of any one of these cationic proteins can damage the membranes of a bacterial cell, causing death by an unknown mechanism. The phagolysosome of neutrophils also contains lactoferrin, an extremely powerful iron-chelating agent that sequesters most of the iron present, potentially inhibiting bacterial growth.
Monocytes and macrophages also secrete a number of the following substances that may play a role in killing pathogens.
- Oxygen metabolites such as O2-, H2 O2, NO and OH. are also secreted into the local environment when the cells bind an Fc receptor on an antibody or when activated by α-interferon.
- Prostaglandins are vasodilators and may increase vascular permeability that accompanies infection and inflammation.
- Monocytes are responsible for the synthesis and secretion of many complement components, which then circulate in the blood and lymph as potent antibacterial factors.
- Stimulation of macrophages by endotoxin causes the synthesis and secretion of interleukin-1 (IL-1). IL-1 along with other cytokines, such as IL-6, stimulates T-lymphocytes to proliferate, thus assisting in the activation of the rest of the adaptive immune system. IL-1 also serves as a chemoattractant for neutrophils and keeps them at the site of inflammation by enhancing the adhesiveness of endothelial cells for neutrophils.
Once a microbe is killed, the phagolysosome employs a large collection of digestive enzymes (e.g., lysozyme, proteases, lipases, nucleases, and glycosylases) that break bacterial macromolecules into low molecular weight components. In neutrophils, spent phagolysosomes accumulate in the cytoplasm and the phagocyte eventually lyses and dies. Dead neutrophils make up most of the material in pus.
Egestion and Antigen Presentation
Macrophages and monocytes live much longer than neutrophils and must dispose of their bacterial components. Once microorganisms are destroyed, the unwanted organic material is expelled from the cell in a process called egestion. Egestion is the opposite of ingestion and the molecular mechanism is basically the reverse of phagocytosis with the microbial leftovers being dumped into the blood and lymph. Some of this microbial debris is not egested, but binds to special protein complexes (called Major Histocompatibility Complex (MHC) molecules) on the membranes of macrophages for presentation to the immune system, as we will discuss when we cover adaptive immunity.
Natural Killer Cells Attack Pathogens, but Are Not Inducible
We end this section with a set of cells that do not fit neatly into either innate or adaptive immunity and these are the natural killer cells. They are a group of nonphagocytic granular lymphocytes present in the body at low populations and containing small amounts of immunoglobulin and MHC I molecules on their surface. Natural killer cells are not T cells, but their behavior is similar to that of cytotoxic T cells. However, natural killer cells are found in animals never exposed to relevant antigens, which suggests that they are not inducible. This is in contrast to T cells that increase in number when exposed to activating antigen. Natural killer cells have Fc receptors on their surface that are capable of binding IgG, which have been shown to trigger antibody-mediated cytotoxicity. While they tend to defy easy classification, it is clear that natural killer cells are responsible for recognizing and destroying virally infected cells, bacteria, fungi, protozoa, helminths and tumors and are, therefore, an important defense against pathogens.
Key Takeaways
- Inflammation occurs in response to tissue damage, either through injury or from infection. In the case of an infection, inflammation causes the activation of the immune response.
- A key part of this immune response is phagocytes that move about the body, are recruited and activated by inflammation, attach to invading pathogens, engulf them, and kill them. Neutrophils, monocytes and macrophages are the three major types of phagocytes.
- Phagocytes detect microbes by the presence of N-formylated peptides, activated complement proteins and the mediators of inflammation. Phagocytes attach to microbes using opsonins, such as IgG and the complement protein C3b.
- In the absence of opsonins, phagocytes can still attach to target microbes nonspecifically, but the process is not as efficient. After binding, phagocytes internalize their prey and fuse them to a lysosome, forming a phagolysosome.
- Once inside the phagolysosome a large collection of oxygen-dependent and -independent mechanisms are used in an attempt to kill the microbe.
- Finally, natural killer cells are part of innate immunity, but respond in a fashion similar to T cells. They attack cells coated with IgG and attempt to kill them.
15 - 7 Summary
A significant amount of our bodies attention is devoted to preventing the invasion of the microbes that are living on us. In this chapter we began our journey through the amazing array of compounds, proteins, cells and organs that are part of our immune system. Every tissue and organ in the body has systems that will prevent and eliminate infection and here we looked at the non-inducible part of the system, innate immunity. The two major arms of innate immunity are inflammation and phagocytes. Inflammation is the warning system that alerts the rest of the immune system that something is wrong, while phagocytes are the infantry of our immune system whose job is to clean out whatever is causing the infection. While innate immunity is powerful, many pathogenic microbes have developed defenses against it. In the next chapter we will look at adaptive immunity and how it combines with innate immunity to defeat almost all infectious agents.
If you enjoyed this free chapter on the immunology, consider buying the book, for only $30.