Latest News
- Clues beginning to emerge on asymtomatic SARS-CoV-2 infection
- Back in November of 2020, during the first wave of the COVID-19 pandemic, I was teaching an in-person microbiology laboratory. One of my students had just been home to see his parents, and they all c…
- Read more
- Could there maybe be better uses of genetics and probiotics?
- Professor Meng Dong and his laboratory have created a probiotic that can metabolize alcohol quickly and maybe prevent some of the adverse effects of alcohol consumption. The scientists cloned a highl…
- Read more
- ChatGPT is not the end of essays in education
- The takeover of AI is upon us! AI can now take all our jobs, is the click-bait premise you hear from the news. While I cannot predict the future, I am dubious that AI will play such a dubious role in…
- Read more
- Fighting infections with infections
- Multi-drug-resistant bacterial infections are becoming more of an issue, with 1.2 million people dying of previously treatable bacterial infections. Scientists are frantically searching for new metho…
- Read more
- A tale of two colleges
- COVID-19 at the University of Wisconsin this fall has been pretty much a non-issue. While we are wearing masks, full in-person teaching is happening on campus. Bars, restaurants, and all other busine…
- Read more
( 52463 Reads)
|Learning Objectives
After reading this section, students will...
- Know that there are many basic similarities between eukaryotic and prokaryotic cell structures, but some structures are different.
- Be able to explain that eukaryotes are the only domain of life to have organelles and why that is true.
- Be able to explain the role of the highly organized cytoskeleton in Eukaryotes.
- Be able to explain how transport is different in eukaryotic cells in comparison to prokaryotes.
- Be able to explain how the structure and function of the nucleus affects transcription and translation.
- Be able to explain how the process of messenger RNA synthesis differs in Bacteria and Eukarya.
This section will focus on describing some of the differences between Bacteria and Eukarya. It will not be an exhaustive review of eukaryotic structure, as you have learned this material in your high school or college biology class.
After the recent journey through the bacterial cell, you may have started to wonder about your cells or other eukaryotic cells. How many properties do we share with bacteria? How are we different? It turns out, as you might expect, we share some basic things in common, but other structures are very different.
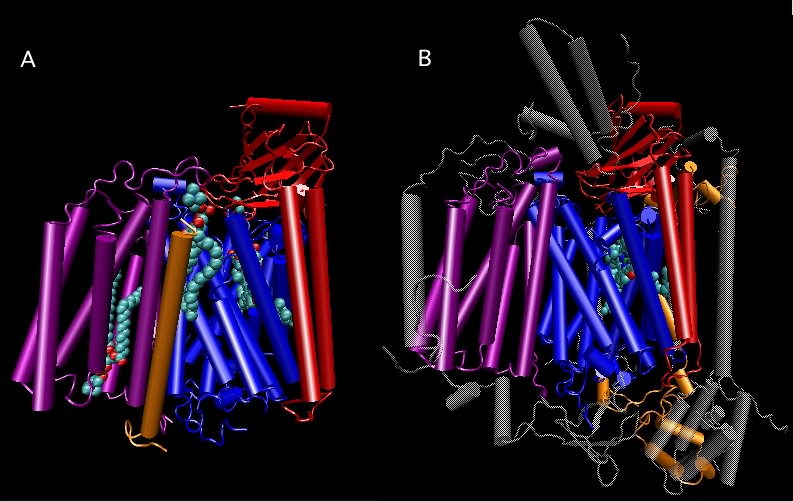
Figure 3.40. Comparison of cytochrome oxidase from bacteria and bovine.. Molecular models of cytochrome oxidase from Rhodobacter sphaeroides (A) and bovine (B) are compared. Each protein is a complex of several distinct proteins, but the four polypeptides shown in color have a high degree of similarity in both their sequence (see Figure 3.41) and structure. The structural similarity should be obvious in this view. Such structurally similarity cannot have arisen by chance, but must reflect the evolution of each from a single ancestor. The gray polypeptides in the bovine cytochrome oxidase are not found in the bacterial protein.
The basic building blocks of the cell, such as nucleic acids, amino acids and sugars are identical. Macromolecular organizations such as chromosomes and membranes have many similarities. Many proteins in eukaryotes, especially those that carry out essential cell functions, have homologs in bacteria that share a high degree of sequence and structural similarity. An example that illustrates this point is the respiratory enzyme cytochrome oxidase. As shown in Figures 3.40 and 3.41, a comparison of cytochrome oxidase from the bovine and Rhodobacter sphaeroides reveals a near-identical arrangement of the catalytic proteins and high sequence similarity. However, the cytochrome oxidase in the bovine has several other polypeptides that serve a structural role.

Figure 3.41. Sequence comparison of cytochrome oxidase from three species. A sequence comparison of cytochrome oxidase showing the high degree of identical amino acids between these very different species: cows and two different bacteria. The colored boxed indicate where the amino acids are identical or similar among the three sequences and the different colors refer to different classes of amino acids.
The DNA in all organisms is chemically similar but the organization of the helix into higher-order structures varies. Eukaryotes contain a larger number of histones (chromosome-binding proteins). In many cases, during cell division, compaction of the chromosome occurs so that they are visible using light microscopy.
The basic mechanism of converting genetic information into proteins is also rather similar. Most of the central components of this process show sequence similarities across biology and substantial functional similarities. The fact that eukaryotes have nuclei means that mRNA must be transported outside of that structure before translation can begin, but the process is otherwise rather similar.
The most conserved of all structures is probably the membrane. Membranes enclose all living systems and every membrane contains amphipathic lipids. The exact chemical structure of the lipids is often different depending upon the species and its environment, but the overall arrangement of the membrane is the same. Membranes perform remarkably similar functions in all species: keeping the cytoplasm in and the environment out. Cholesterol is common in the membranes of eukaryotes, but is uncommon in bacterial and archaeal membranes.
Things that are different between eukaryotes and prokaryotes
Eukaryotes are typically more complex than prokaryotes and appear more organized when examined in the microscope. This organization into different intracellular compartments likely reflects the demands of a more complex cellular system. It may also be that the formation of these separate organelles allowed the subsequent evolution of more elaborate cells. Figure 3.42 diagrams the various structures found in the typical eukaryotic cells. Only photosynthetic organisms contain chloroplasts. The organelles are the nucleus, the mitochondria, the endoplasmic reticulum and the Golgi apparatus. Internally eukaryotic cells have a cytoskeleton that determines cell structure, and plants supplement this with cell walls. We discuss these structures and their functions in eukaryotic cells in the following sections.
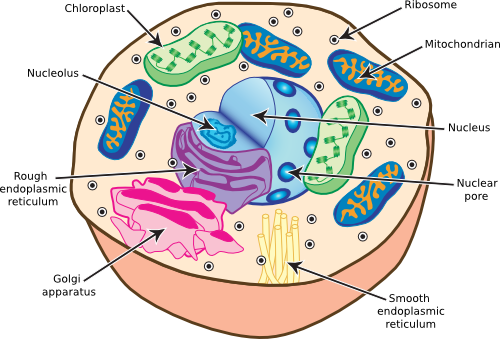
Figure 3.42. Eukaryotic cell structure. A diagram of the common structures found in eukaryotic cells.
Microfilaments and microtubules
The cytoskeleton is a network of filaments and fibers found in the cytoplasm of many eukaryotic cells. It serves four known roles in cells.
- The cytoskeleton helps to determine the shape of the cell.
- It is involved in cell division, allowing the separation of the chromosomes into daughter cells.
- The cytoskeleton moves organelles around the cell.
- It is involved in motility either through the use of flagella or by amoeboid movement.
The cytoskeleton components can be divided into three classes based upon the filaments' size, distribution, and function. Microfilaments, made from actin, are the smallest at 4 to 6 nm in diameter. These lie beneath the surface of the cell membrane and are anchored to it, forming a web-like structure. They dictate the cell's shape and can also be involved in motility by contraction or expansion of the filament. Filaments may also tether organelles to the membrane and help move them around the cell. This movement can be important for the modulation of organelle function. Intermediate filaments are 10 nm in diameter and made of keratin, the same protein found in hair and fingernails. These filaments take different forms and are present in many types of cells, but their exact function is unknown. They may play a structural role similar to some microfilaments. Figure 3.43 shows some examples of cytoskeletal elements
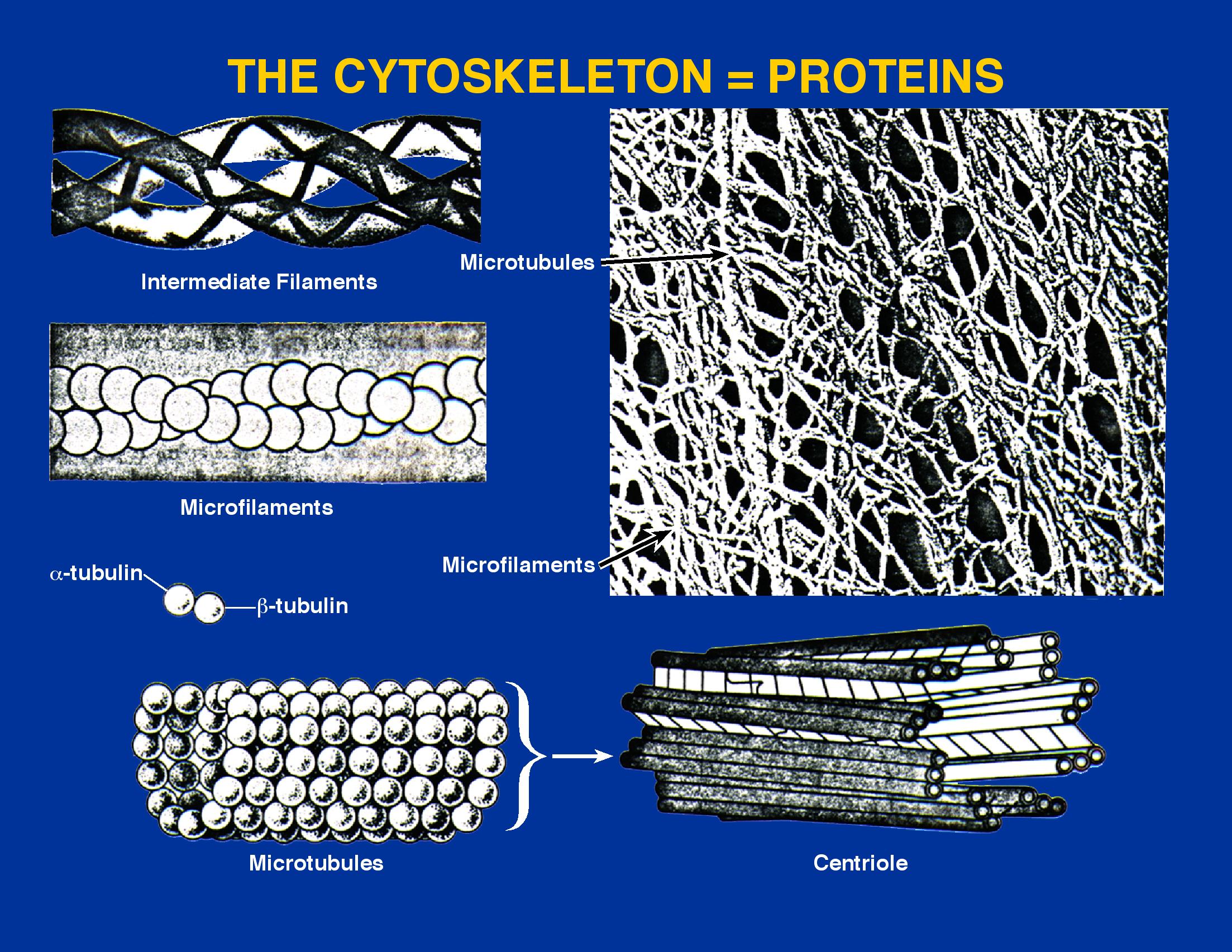
Figure 3.43. Cytoskeletal elements. Eukaryotic cells have several different types of scaffolding proteins to help them keep their shape. Microfilaments, microtubules, and centrioles are all important structural elements.
In addition to the above, there are more complex fibers and structures. Microtubules are hollow cylindrical structures 20-25 nm in diameter containing tubulin as the major structural protein. Tubulin polymerizes into a helical cylindrical structure, and thirteen of these protofilaments then combine to make a microtubule. Microtubules can form the basis of many different structures. Many of these structures form to perform a necessary function and are then disassemble. Often microtubules form centrioles that contain nine sets of microtubules arranged in a circular matrix. These are 4.0 µm long and 1.50 µm wide and are usually found in pairs at right angles to each other. Centrioles are important in proper chromosome segregation and cell division, as discussed in the section on the nucleus below. Microtubules are also part of the basal bodies of flagella and cilia.
Cilia and flagella are examples of more permanent structures that contain microtubules. Both cilia and flagella have a similar structure of nine pairs of microtubules arranged in a circular fashion around a tenth pair that runs down the center. Both are attached to the membrane and project into the environment. In a process that requires energy, the microtubules in the outer ring are moved with respect to each other, causing the cilia or flagella to bend and snap back in a whip-like fashion. This bending causes the movement of liquid near the structures such that spent liquid with few nutrients and waste products is moved away from the cell and replaced by fresh liquid containing nutrients and oxygen. The beating of cilia and flagella can also push the cell through its environment. Cilia are 2 to 10 µm long and 0.5 µm wide and are shorter and typically more numerous than flagella, with hundreds of them on some types of cells. An example of a ciliated organism is the unicellular protist Paramecium found in freshwater ponds. The microbe is a predator of bacteria, and motility is vital in this lifestyle, both for chasing down prey and moving away from danger. Cilia cover the surface of Parameciumand move the organism through the environment by beating in a coordinated fashion.
Flagella are 5-100 µm in length, and there are typically only one or two per cell. Eukaryotic flagella are larger than those found on bacteria or archaea and have a more complex structure. Flagella are found in many unicellular creatures, and their primary role is cell motility.
Endoplasmic reticulum
The endoplasmic reticulum (ER) is a finely divided system of interconnected membranes, consisting of tubules and vesicles that loop through the cell and are contiguous with the nuclear membrane. A drawing of the ER is shown in Figure 3.44. It functions in the synthesis of membranes and membrane proteins and is also involved in protein secretion. Not surprisingly, the ER is especially prominent in cells doing a large amount of protein secretion. The ER works very closely with the Golgi apparatus (see below) to carry out these functions. No structure in bacterial cells is analogous to the ER, but many of the same functions occur on the inside surface of the cellular membrane in bacteria. ER comes in two types: rough ER and smooth ER.
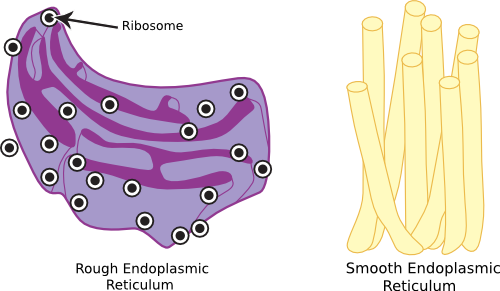
Figure 3.44. The endosplasmic reticulum. Eukaryotic cells contain a network of passages that connect various elements of the cell and are also important in secretion.
Rough ER gets its appearance from the presence of ribosomes on its surface, as seen in the electron microscope, and its function is the production, processing, and export of proteins. During translation, an appropriate signal guides the ribosome to the ER, causing protein synthesis directly across the membrane into the lumen. These proteins may be processed or modified by the addition of carbohydrates to form glycoproteins. After processing, proteins move slowly through the ER and are packaged into vesicles of ER membrane called transition vesicles. These release from the ends of the ER and move via the cytoskeleton to either to the Golgi apparatus or to the plasma membrane. Once the transition vesicle contacts the Golgi (or plasma membrane), the two fuse and release the contents of the vesicle into the target compartment.
Smooth ER does not contain ribosomes, and the lumen and membrane of smooth ER contain a variety of enzymes that perform many functions, including modification of toxins and synthesis of steroids.
Golgi apparatus
The Golgi apparatus is an organelle containing a double membrane, and it is mainly devoted to the processing of proteins synthesized in the ER. A drawing of the Golgi apparatus is shown in Figure 3.45. It is found in many eukaryotic cells, but it lacks a well-formed structure in many fungi and ciliate protozoa. It consists of regions of stacked contiguous membranes containing no ribosomes. Each membrane sac is 15 to 20 µm thick and separated from the next stack by about 30 µm. A complex network of tubes and vesicles extend from the edges of these sacs into the surrounding cytoplasm. The Golgi has a definite polarity with membranes near the ER (the cis face) having a different shape and enzyme content than those at the opposite end (the trans or maturing face). Studies of the Golgi apparatus appear to show material flowing into the cis face from vesicles through the apparatus and then exiting at the trans face.

Figure 3.45. The Golgi Apparatus. The Golgi apparatus is involved in the glycosylation and proteolytic processing of proteins that are to be secreted into various cellular organelles or to the outside environment. Proteins often pass through the Golgi apparatus as part of their maturation
The role of the Golgi apparatus is to package material for export, but its exact function varies depending upon the organism. For example, Giardia and Entamoebautilize Golgi apparatus to form cell walls during cyst formation. It often participates in the synthesis of cell membranes and the final processing of proteins before export. In many cases, the Golgi apparatus contains glycosylation enzymes that add sugars to proteins as they move through its lumen. The type of glycosylation that takes place is dependent upon signals contained within the protein sequence. The Golgi also processes enzymes using proteases, which clip at specific amino acids sequences to form mature proteins and hormones. These mature proteins then move to their final destinations, which may be in the membrane, in lysosomes or secreted into the environment.
Lysosomes
One of the most important functions of the Golgi apparatus is the synthesis of lysosomes, an organelle that is present in a variety of eukaryotic cells. Lysosomes are spherical structures enclosed in a single membrane that can vary in size from 50 nm to several µm. They are involved in intracellular digestion and contain enzymes (called hydrolases) that digest many types of macromolecules. Hydrolases function best under acidic conditions (pH 3.5 to 5.0) and the lysosome maintains this pH by membrane proteins that pump protons into its interior. Ribosomes synthesize enzymes bound for lysosomes on the ER. They then move through the smooth ER and Golgi apparatus before being package into lysosomes. In some cases, lysosomes can also bud off from the smooth ER.
Lysosomes serve a variety of functions depending upon the cell type. In unicellular eukaryotes, they are often digestive structures that take bacteria or other substances from the outside environment and degrade them into usable nutrients. In mammals, lysosomes serve to eliminate unwanted particles, either cell structures that are no longer needed or foreign macromolecules (from viruses or bacteria) that have invaded the cell. In any case, the hydrolytic enzymes and low pH typically inactivate and then degrade any particle that enters the lysosome
Eukaryotic cells absorb things by endocytosis
Lysosomes are an important part of endocytosis, a process that eukaryotic cells use to take up particles from the environment. Figure 3.46 illustrates endocytosis. The process creates membrane-bound cavities filled with fluid and solid materials. Larger membrane-enclosed cavities are called vacuoles, while smaller ones are called vesicles. Endocytosis comes in two forms, phagocytosis and pinocytosis. Phagocytosis involves the engulfment of large particles, even microorganisms, into membrane-bound compartments. It is a process used most often in the immune system, and the chapter on the innate immune system describes it in detail. Pinocytosis involves the recognition of specific particles in the environment, as described below. Unicellular eukaryotic microbes use the process to ingest food, while multicellular organisms use it to take in certain macromolecules traveling from other parts of the organism.
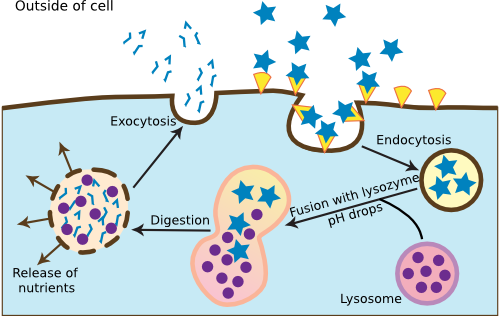
Figure 3.46. The process of endocytosis. Eukaryotic cells absorb materials from the outside by using endocytosis. This is a standard pathway through which most material enters the cell.
Pinocytosis begins when protein receptors on the cell surface bind to the target molecule to be ingested. The bound particle migrates to an area of the membrane that is rich in a protein called clathrin. This protein forms a matrix and causes an indentation in the membrane called a clathrin-coated pit. The entrance of a receptor:particle complex begins the invagination process at the pit that results in the engulfment of the incoming particle inside a membrane vesicle. This vesicle is called an endosome and once inside the cell, proteins in the endosome membrane begin to pump protons inside, dropping the internal pH of the endosome. The clathrin on the endosome membrane then migrates back to the plasma membrane to repeat the cycle. The endosome fuses with a lysosome to form an endolysosome, which causes the degradation of the ingested particle. The cell then transports the valuable breakdown products to their destinations. The spent endolysosome is called a residual body and sometimes fuses with the plasma membrane to release remaining compounds into the environment.
The ER, Golgi apparatus, lysosomes, and endosomes seem to operate as a coordinated whole, functioning to import and export materials. It is probably correct to think of these structures as a functional unit, and the term vacuome has been coined to describe them. Ribosomes in the ER manufacture proteins and the ER and Golgi modify them. The mature proteins eventually find their way to the plasma membrane for export or become part of lysosomes. Lysosomes serve in the endocytotic pathway to take up materials and process them for cell use. They also digest spent cell constituents. All of these processes occur within membrane structures. This carefully controls the import and export of materials from the cell. Considering the extent of these structures in the cell, it is remarkable that the membranes of these structures, especially lysosomes, never rupture. If they did, it would be catastrophic to the cell and would rapidly lead to cell death.
The nucleus holds the cell's genetic material in eukaryotes
While the bacterial cell does seem to sequester its chromosome to a portion of the cytoplasm, no demarcation divides the nucleoid from the rest of the cell. In eukaryotes, the nuclear membrane separates the cell's DNA from the cytoplasm. The nucleus is the largest and most visible organelle of eukaryotic cells. It contains almost all the cell's DNA and is the site of chromosome replication and transcription. It has two layers of membrane encircling it called the nuclear envelope, with the outer layer being contiguous with the ER. Scattered throughout this nuclear envelope are circular openings known as nuclear pores. These pores are highly discriminatory, allowing easy movement in and out of the nucleus of only appropriate macromolecules such as proteins with specific sequences.
In eukaryotes, the chromosome is not a single circular piece of DNA like most prokaryotes. Rather, most eukaryotic cells split their genomes into several linear chromosomes, with each cell containing at least two copies of each chromosome. The exceptions are those cells specializing in reproduction and only contain one copy of the cell's chromosomes. Each piece of DNA is complexed with special basic structural proteins called histones that seem to be important in keeping the DNA organized. Proteins bind the DNA, and the entire set of DNA and associated proteins is called chromatin. For much of the cell cycle, chromatin consists of long DNA strands formed into beads by the association of histones along its length. Figure 3.47 shows a nucleus during mitosis, with the chromosomes visible.
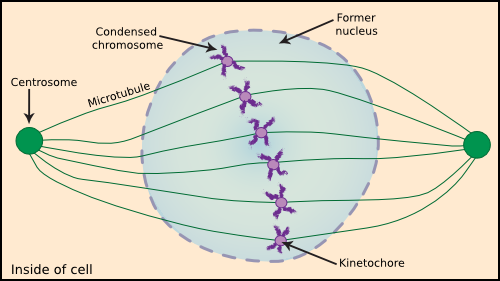
Figure 3.47. A eukaryotic cell in the middle of mitosis. Eukaryotic DNA replication takes place during the cell phase called mitosis. At this time, protein synthesis is halted and the chromosomes condense. The sister chromatids meet at the middle of the cell and then migrate to two separate poles. This movement is coordinated by centrosomes, kinetochores and microtubules.
In actively growing cells, the DNA is replicated from numerous sites rather than the single bi-directional origin in prokaryotes. These multiple DNA origins are necessary due to the much larger amount of DNA found in most eukaryotic cells. During division in prokaryotes, the cell appears to simply split in two, with each daughter cell receiving a chromosome. In contrast, eukaryotic cells go through a morphologically distinct phase, mitosis, to separate the chromosomes. One of the more important events of mitosis is the binding of additional histones and the contraction of the chromatin into compact structures that were called chromosomes due to their staining properties. (The original meaning of the term chromosomes is a colored body, but is now synonymous with a cell's DNA). The two daughter chromosomes formed during replication are physically separated into the separate daughter cells by the filaments called microtubules. These attach at one end to the chromosome at a region termed the kinetochores, and at the other end, they attach to one of two regions of the cell called a centrosome. By depolymerizing the microtubules at each centrosome, each daughter chromosome is pulled away from its partner and toward a region that eventually reforms as a new nucleus.
There are also some important differences in transcription between eukaryotes and prokaryotes. In eukaryotes, mRNA transcription takes place in the nucleus, and the finished mRNA moves through the nuclear pores and into the cytoplasm for translation by the ribosomes. The genes of eukaryotic cells also contain regions of largely unimportant DNA, termed introns, that do not code for protein. After the transcription of a gene into mRNA, enzymes remove these introns before translation. One set of nuclear proteins removes these sequences and splices the actual coding sequences (exons) together to make the finished mRNA. The finished mRNA then travels from the nucleus to ribosomes in the cytoplasm. Modifications of both ends stabilize the mRNA of eukaryotic cells. At the front end is usually a 5'-cap made of 7-methylguanine attached to the mRNA by a triphosphate linkage. At the 3' end of the mRNA is a long stretch of A bases (a poly-A tail) that have a role in mRNA stability, as mentioned earlier in this chapter. Finally, while it is quite common in bacteria to have several genes on each mRNA transcript, the vast majority of mRNAs in eukaryotes code for only a single protein product. Figure 3.48 shows the steps in gene expression in eukaryotes.
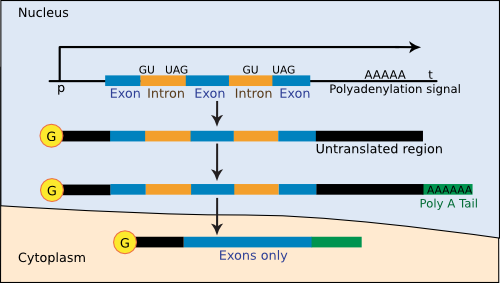
Figure 3.48. The steps of gene expression in eukaryotes. Processing genetic information into protein is more complex in eukaryotes than in prokaryotes. In part this is due to the existence of introns in most genes of "higher eukaryotes" (though introns are rare in yeast). These have to be removed, a poly-A tail added to the 3' end and a cap added to the 5' end of the mRNA before it reaches the cytoplasm for translation.
Eukaryotic cells also contain one or more dark-staining structures within the nucleus called nucleoli. Although they are not enclosed within a separate membrane, the nucleoli are complexes with separate granular and fibrillar regions. They are present in non-dividing cells but frequently disappear during mitosis and reappear after cell division. The nucleolus is the initial site of ribosome synthesis. This structure contains the DNA that codes for the ribosomal RNA genes. The ribosomal RNAs are synthesized and then processed to form the final rRNA molecules. These then combine with several ribosomal proteins synthesized in the cytoplasm to form an initial ribosomal complex. The entire complex then migrates out of the nucleus into the cytoplasm, where it combines with other proteins to form a ribosome.
Mitochondria and plastids are organelles of energy generation in eukaryotic cells
Mitochondria are involved in energy generation through respiration. Mitochondria have no fixed shape but often look like short rods in transmission electron micrographs when viewed along their long axis (Figure 3.49). Each mitochondrion contains two membranes. The outer membrane is smooth and serves as a selective barrier. The inner membrane is highly convoluted and folded and contains a high number of membrane complexes. Nutrients are oxidized inside the mitochondria by catabolic enzymes, and the high-energy electrons extracted are donated to a respiratory chain in the inner membrane. These enzymes then create a proton gradient, and this gradient is then used to synthesize ATP. ATP leaves the mitochondria, and it serves as a source of energy for the rest of the cell's machinery.
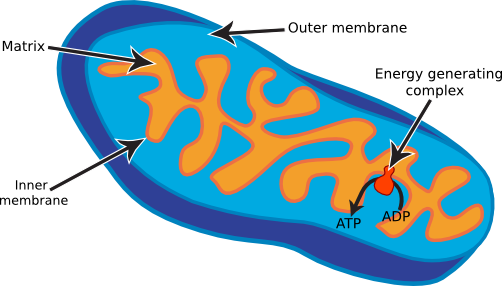
Figure 3.49. Mitochondria structure. Mitochondria are rod-shaped structures that resemble the shape of common bacteria. They contain two membranes, similar to what is found in gram-negative bacteria, and 70S ribosomes. Energy generation occurs at the inner membrane.
Plastids are specialized organelles involved in metabolism that are unique to plants and come in several forms. Amyloplasts are starch storage containers found in some plants. Chloroplasts are oval-shaped structures inside plant and algal cells containing an outer and inner membrane, as shown in Figure 3.50. The outer membrane serves a similar function to the outer membrane of mitochondria, while the inner membrane consists of a network of stacks of membranous disks, called thylakoids, which are attached together by narrow tubes of membrane. The thylakoid membranes are the centers of photosynthesis in eukaryotes. They contain enzyme complexes that capture light and produce ATP and high-energy electrons. These then power the synthesis of organic compounds from carbon dioxide.
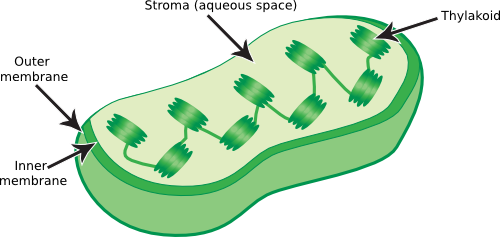
Figure 3.50. Chloroplast structure. Chloroplasts are the site of the light reactions of photosynthesis. Light striking the chloroplast is converted into a proton motive force and this is used to generate energy. Chloroplasts contain two membranes, again similar to gram-negative bacteria, and the chloroplast itself is a relative of cyanobacteria.
Mitochondria and chloroplasts each have a single circular chromosome and large numbers of ribosomes that are of bacterial (70S) and not eukaryotic (80S) form. The presence of two lipid bilayers, a circular chromosome, and 70S ribosomes is consistent with the evolutionary hypothesis that we have already explained near the beginning of this chapter.
The implications of eukaryotic structures on their growth
Eukaryotic cells are generally much larger than prokaryotic ones, and this difference in volume has several implications. First bigger cells can afford to have more things stored in the cytoplasm. This larger size means it is not as costly for a eukaryotic cell to have structures taking up space. In a prokaryote, space is at a premium, and the cell rapidly degrades anything not being used. This larger size may be one reason that organelles are possible. Second, larger cells have a lower surface-to-volume ratio than smaller cells, and therefore, prokaryotes effectively have more contact with their environment. This greater exposure can mean a more rapid response to changing environmental conditions. Finally, bigger cells have more of a challenge moving molecules within themselves. Prokaryotes can often depend on simple diffusion to move molecules around the cell, but this process might be too slow and inefficient in much larger cells. Eukaryotes overcome this by having specific transport mechanisms (i.e., microtubules) inside the cell.
Size constrains eukaryotes in two important ways: how fast they grow and what environments they can tolerate. The compartmentalization of the genome inside the nucleus limits the rate at which eukaryotic cells can divide. The complete cell division cycle in a multicellular eukaryotic organism depends on the cell type, but it takes at least 8 hours even in rapidly dividing skin cells. In unicellular yeast cultures, the shortest cell cycle is about 1.7 hours under ideal conditions. Due to their smaller genomes, lack of a nucleus, and the ability to couple transcription and translation, bacteria can grow much faster. Clostridium perfringens has been shown to go through a complete division cycle in at little as 6.6 minutes at 43 °C in beef cubes!
There are environments that have one or more physical characteristics that prevent the growth of most organisms. These extreme environments might be too hot, cold, acidic or alkaline for typical organisms to grow. However, a small subset of prokaryotes has evolved to take advantage of these environments and thrive, and prokaryotes always define the extremes of where life can exist. In part, this is probably because simpler cells have fewer "body parts" that must be changed for growth under very different conditions. In eukaryotes, they would need to modify their cytoplasmic contents to tolerate the extreme environment and the makeup of their organelles.
Key Takeaways
- At the molecular level, Bacteria and humans are more alike than different. However, these differences are important.
- First eukaryotes are larger, and this allows them to carry more DNA and other structures. The DNA can therefore code for more genes, enabling the control and regulation that is necessary in a multicellular organism.
- This complexity is manifested in the various visible structures of the cell, including: mitochondria (energy generation), the nucleus (DNA storage and transcription), chloroplasts (photosynthesis), the Golgi apparatus, the endoplasmic reticulum, lysosomes (the last three involved in protein export).
- The larger size of Eukarya cells necessitates the presence of a larger cytoskeleton, consisting of microfilaments and microtubules, involved in the transport of materials about the cell and the formation of structural components.
- The increased complexity of eukaryotes also makes eukaryotic cells less adaptable and unable to survive in extreme environments. Their large size slows the production of proteins, and eukaryotes grow more slowly than prokaryotes.
Quickcheck 3-9
Warning, you must be logged in to be able to have your exam graded. Answer the questions below and if you are a registered user of the site you will see a Grade Exam button. Click it to have your exam graded.