Latest News
- Clues beginning to emerge on asymtomatic SARS-CoV-2 infection
- Back in November of 2020, during the first wave of the COVID-19 pandemic, I was teaching an in-person microbiology laboratory. One of my students had just been home to see his parents, and they all c…
- Read more
- Could there maybe be better uses of genetics and probiotics?
- Professor Meng Dong and his laboratory have created a probiotic that can metabolize alcohol quickly and maybe prevent some of the adverse effects of alcohol consumption. The scientists cloned a highl…
- Read more
- ChatGPT is not the end of essays in education
- The takeover of AI is upon us! AI can now take all our jobs, is the click-bait premise you hear from the news. While I cannot predict the future, I am dubious that AI will play such a dubious role in…
- Read more
- Fighting infections with infections
- Multi-drug-resistant bacterial infections are becoming more of an issue, with 1.2 million people dying of previously treatable bacterial infections. Scientists are frantically searching for new metho…
- Read more
- A tale of two colleges
- COVID-19 at the University of Wisconsin this fall has been pretty much a non-issue. While we are wearing masks, full in-person teaching is happening on campus. Bars, restaurants, and all other busine…
- Read more
( 47766 Reads)
|Learning Objectives
After reading this section, students will...
- Be able to explain the structure and function of resting cells.
- Be able to explain why endospores are resistant to heat, chemicals and radiation.
- Be aware of different cells states into which microbes can differentiate.
For obvious reasons, we have focused on growing cells, but there are non-growing states of microbes that are important to both microbes and humans. In these states, termed spores and cysts, the cells remain dormant for long periods of time. Part of the relevance of these states is that the same properties that allow the cell to survive extended periods also happen to make the cells resistant to our typical efforts to kill them. Consequently, spores and cysts can thwart attempts to sterilize a sample. In this section, we examine some of the properties of these structures.
Spores and cysts
Spores and cysts are resting structures. That is, these states have very low to nonexistent rates of metabolism. They are common in organisms that live in soil and may need to survive some rough conditions such as lack of nutrients, high heat, radiation, or drying.
Sporulation is a unique developmental cycle. After a cell decides to sporulate, the creation of a different type of cell needs to take place, which requires turning on a large collection of genes in a tightly coordinated fashion. In addition, all of this expression must be complete before the microbe runs out of energy. There are several types of spores. Some are highly resistant structures that form under conditions of cell stress inside a supportive cell. These are called endospores. Others are part of the normal reproductive cycle, being created by the differentiation of a vegetative cell, and we will refer to these as spores. In this section, we talk generally about the structure of spores, and in the chapter on regulation, we will examine the regulation of sporulation.
Endospores
Endospores are refractile - light cannot penetrate them - so that they are easy to see in the phase microscope, and this makes it easy to detect them. Most endospores have no measurable metabolism and are a form of suspended animation. Endospores can survive for a very long time and then return to a growing state, a process termed germination. Endospores that were dormant for thousands of years in the great tombs of the Egyptian Pharaohs were able to germinate and grow when placed in appropriate medium. Several scientists have recovered viable endospores from bees trapped in amber that is 25-40 million years old. The microbe isolated was found to be most closely related to Bacillus sphaericus. There are even claims of endospores over 250 million years old being able to germinate when placed in an appropriate medium, but these claims still need to be verified. Endospores are everywhere, are easily dispersed throughout the environment, and can be difficult to remove. The anthrax scare of 2001 in the United States is ample evidence of the insidiousness of endospores and their impressive resistance.
The formation of an endospore is clearly a great advantage for these bacteria and enables them to endure extreme stress. At a later time, even much later, when conditions are favorable, they can reemerge and flourish. Endospores enable a species to spread easily from one suitable environment to another, and many endospore-forming bacteria are ubiquitous in the environment. Endospores are a particular problem in the food industry. Companies must take great care to ensure either the destruction of endospores or suitable preservation methods so that endospore-forming bacteria (and other microbes) cannot grow.
Endospores are resistant to heat (>100 °C), radiation, many chemicals (i.e. acids, bases, alcohol, chloroform), and desiccation. The mechanisms that account for this resistance include the impermeability of the endospore coat, the dehydration of the cytoplasm and the production of special proteins that protect the spore's DNA. Figure 3.34 shows the major structures of an endospore.
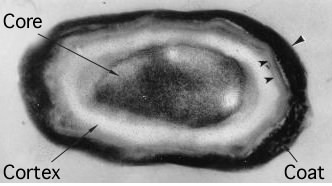
Figure 3.34. An endospore. An electron micrograph of an endospore of Bacillus subtilis showing the core, cortex and coat. (Source: M. Serrano, et al. 1999. J. Bacteriol. 181:3632-3643)
Endospores can be divided into several important parts (Fig 3.51). The center of the endospores contains the core, and it consists of the cytoplasm, DNA, ribosomes, enzymes, and everything needed to function once returned to the vegetative state. The core is dehydrated, which is essential for heat resistance, long-term dormancy, and maximum chemical resistance. Calcium dipicolinate is a major component of the core and plays a role in resistance to wet heat and UV light. The cortex surrounds the core and comprises two layers, a thin, more densely staining layer that is similar in structure to the vegetative cell wall and a thicker, less dense layer containing modified peptidoglycan. Two major modifications are present. First, there is less cross-linking with only 3% of the muramic acid present in the peptidoglycan of the cortex participating compared to 40% of muramic acid in the vegetative cell wall. Second, much of the muramic acid is modified to a muramic-β-lactam structure. Both of these modifications of the cortex appear to be important in germination. Muramic-β-lactam serves as a specific target for lytic enzymes that activate during germination, and the lower cross-linking enables easier outgrowth. Outside of the cortex is the spore coat composed of more than two dozen different types of proteins. There is some evidence that cross-links connect these proteins. This covalent connection between coat proteins probably contributes to the spores' resistance.
Some microbes make other types of resting structures
Endospores are not the only type of spore made by microorganisms. Azotobacterspecies and many others are known to form cysts, which are dormant cells with thickened cells walls (Figure 3.35). Cysts are resistant to desiccation and some chemicals, but cannot withstand high temperatures as endospores can.
The actinomycetes are a large group of spore-forming, gram-positive bacteria that grow by forming long tubules called filaments. Under nutrient-poor conditions, these filaments differentiate into round resting structures termed spores. In contrast to endospores and cysts, these structures are part of the reproductive process. The developmental process to create an actinomycete spore is less complex than that of the endospore. It involves the simple formation of cross walls that divide the filament into sections, each containing a chromosome. These then differentiate into mature spores. During this process, the bacterium lays down a tougher cell wall, and there is a conversion of the cytoplasm to a dormant state so that the spore becomes more resistant to heat and chemicals, though not as hardy as an endospore. Actinomycete spores can survive for long periods (for years) and germinate into vegetative cells when appropriate growth conditions are present. Many different genera are capable of forming this type of spore, and the ability to form these structures does not seem to correlate with any group of microorganisms.
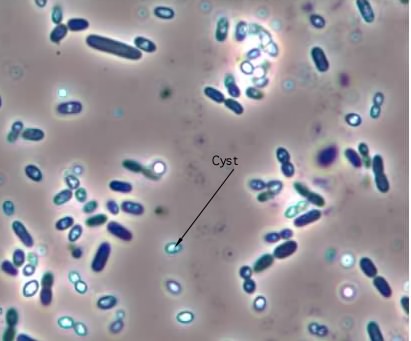
Figure 3.35. A cyst. The micrograph shows cysts and cells of Azotobacter vinelandii at 1000 X magnification. The cysts are the phase-bright objects, while the cells are darker.
Heterocysts are differentiated cells that specialize in nitrogen fixation
Some photosynthetic filamentous cyanobacteria are capable of forming specialized structures called heterocysts. These are rounded structures distributed at regular intervals along the string of vegetative cells, as shown in Figure 3.36 or at one end. Heterocysts evolved to solve the problem of performing plant-like photosynthesis (which produces oxygen) and at the same time fixing N2to ammonia (a process that involves enzymes that inactivate in the presence of oxygen). The single focus of the heterocysts is to fix N2, while the rest of the cells perform photosynthesis (and divide), thus keeping the two processes separate. Heterocysts develop a surface that is impermeable to gasses, and begin synthesizing large amounts of nitrogenase, the protein that fixes N2. Importantly, these heterocysts maintain some permeability to the cells on either side. Neighboring cells take N2from the atmosphere and pass it along to the heterocyst, which reduces it to NH3and returns fixed nitrogen to its neighbors. The neighboring cells also prevent oxygen from reaching the heterocyst. Heterocysts are essentially specialized organs for the "multi-cellular organism" represented by a chain of cyanobacterial cells that only form when nitrogen is limiting. The regulation of this developmental cycle is intriguing and serves as a simple example of multicellular development in a unicellular organism.
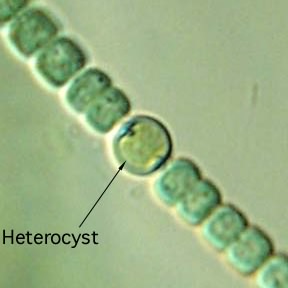
Figure 3.36. A heterocyst of a filamentous cyanobacteria from the Anabaena genus. Heterocysts are common in several different groups of cyanobacteria and are the site of nitrogen fixation. Note the slightly enlarged size and distinct shape of the heterocyst when compared to the vegetative cells on either side. (Source: Michael Clayton, University of Wisconsin-Madison)
This ends our survey of the cellular structure of bacteria. In the remaining sections, we look at the major structural differences that distinguish the archaea and eukaryotes from the bacteria.
Key Takeaways
- Cells can also differentiate to form visible structures that are not vegetative cells. Spores and cysts are resting structures that are a form of suspended animation.
- These structures allow a cell to avoid environmental challenges and germinate at a later time when conditions are better.
- Endospores can survive for very long periods and survive very harsh conditions.
- Heterocysts are another structure that cyanobacteria form to help them fix nitrogen. Heterocysts allow cyanobacteria to fix nitrogen, an anaerobic process, at the same time that they are performing photosynthesis, a process that generates oxygen.
Quickcheck 3-7
Warning, you must be logged in to be able to have your exam graded. Answer the questions below and if you are a registered user of the site you will see a Grade Exam button. Click it to have your exam graded.