Latest News
- Clues beginning to emerge on asymtomatic SARS-CoV-2 infection
- Back in November of 2020, during the first wave of the COVID-19 pandemic, I was teaching an in-person microbiology laboratory. One of my students had just been home to see his parents, and they all c…
- Read more
- Could there maybe be better uses of genetics and probiotics?
- Professor Meng Dong and his laboratory have created a probiotic that can metabolize alcohol quickly and maybe prevent some of the adverse effects of alcohol consumption. The scientists cloned a highl…
- Read more
- ChatGPT is not the end of essays in education
- The takeover of AI is upon us! AI can now take all our jobs, is the click-bait premise you hear from the news. While I cannot predict the future, I am dubious that AI will play such a dubious role in…
- Read more
- Fighting infections with infections
- Multi-drug-resistant bacterial infections are becoming more of an issue, with 1.2 million people dying of previously treatable bacterial infections. Scientists are frantically searching for new metho…
- Read more
- A tale of two colleges
- COVID-19 at the University of Wisconsin this fall has been pretty much a non-issue. While we are wearing masks, full in-person teaching is happening on campus. Bars, restaurants, and all other busine…
- Read more
( 45706 Reads)
|Learning Objectives
After reading this section, students will...
- Know the structure and functions of flagella, pili and fimbriae.
- Be able to explain the composition and purpose of capsules and slime layers.
Surface structures are typically attached to a membrane, and extend into the environment. Important structures include flagella, pili, fimbriae, and glycocalyx. These protrusions and surfaces interact with the environment around the microorganism and therefore are pivotal in how the microbe sees the world and how we see the microbe.
Flagella are one type of structure used for motility
In many bacteria, flagella are responsible for motility in liquid. There is also a loose correlation between cell shape and the presence of flagella. Almost all spirilla, half of all rod-shaped bacteria, and only a few of the cocci are motile by flagella. One rationale for this correlation might be that spherical cells such as the cocci simply do not have the best geometry for directional movement by flagella, unlike more linear bacteria.
Flagella can be thought of as little semi-rigid propellers free at one end and attached to a cell at the other. Flagella are thin (20 µm) and long, with some having a length 2-3 times (about 10 µm) the length of the cell. Due to their small diameter, flagella cannot be seen in the light microscope unless a special stain is applied. Bacteria can have one or more flagella arranged in clumps or spread over the cell surface. Figure 3.29 demonstrates some of the more common arrangements.

Figure 3.29. Flagellar arrangements. A cartoon of several common flagellar arrangements.
Flagella are mostly composed of the protein flagellin, which is bound in long chains and wraps around itself in a left-handed helix as shown in Figure 3.30.
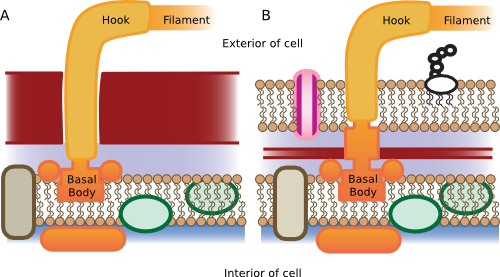
Figure 3.30. Flagella structure. Flagella are attached by a hook and rings that anchor it to the cell wall of the microorganism. In gram-positive bacteria (A) the rings are located in the cytoplasmic membrane and the flagella passes through the peptidoglycan to the outside environment. In gram-negative bacteria (B) there are additional protein rings in the outer membrane.
Flagella are organs of motility
The flagellum attaches to the cell through complex protein structures termed the hook and the basal body, which span the cell membrane. One ring in the basal body rotates relative to the other, causing the flagellum to rotate. The proton gradient, created by metabolism, provides the energy to drive this basal body (we will discuss this in much more detail in the metabolism chapter). In some fashion, the translocation of protons from outside to inside the membrane causes the rotation of the flagellum. In a sense, protons move through the wheel-like basal body (similar to a water wheel, except using protons), which causes the rotation of the assembly, including the flagellum. When E. coli is swimming through a solution, the flagella turn counter-clockwise and push the microbe. This behavior is termed smooth swimming. It is possible for E. coli to also reverse the direction of flagellar rotation, and when the flagella turn clockwise, they pull against the bacterial cell. Since E. coliis flagellated peritrichously (that is, at many positions), it is pulled in all directions and tumbles.
How fast do bacterial cells move? The average is 50 µm/sec, which is about 0.00015 kilometers/hr. speed may seem slow but remember their tiny size. Table 2.4 shows a better comparison and indicates that, relatively speaking, bacteria are faster than humans. Also, remember that this motility happens in water, which is much more viscous than air.
Table 2.4 Relative speeds of organisms
| Organism | Kilometers per hour | Body lengths per second |
| Cheetah | 111 | 25 |
| Human | 37.5 | 5.4 |
| Bacteria | 0.00015 | 10 |
Bacteria seem slow, until you consider relative size. Then they are quite fast.
If a flagellum breaks off, it is resynthesized until it reaches the appropriate length. This growth occurs from the tip. The flagellar filament is hollow, and flagellin monomers are passed up through this space until they reach the growing tip and are addadded to the structure.
Taxis and flagella
Typically microbes in aqueous environments continually move around looking for nutrients. Even microorganisms in the soil have uses and opportunities for movement. Sometimes this movement is random, but in other cases, it is directed toward or away from something. As a rough guide, bacteria want to move towards food or energy sources and away from toxic compounds. In other words, bacteria are capable of showing simple behavior that depends upon various stimuli.
There are several classifications of tactic responses, and the category is based upon the stimulus that the movement is responding to.
- chemotaxis - towards or away from a chemical stimulus
- phototaxis - towards light
- aerotaxis - towards or away from oxygen
The purpose of chemotaxis should be fairly evident: attraction to nutrients or avoidance of damaging compounds. A cell distinguishes chemicals that fall into either of these classes with receptor molecules near their surfaces that can tell if specific chemicals are in the environment. Many of these receptors are also involved in transport of their target molecules. Now it is trickier than it sounds. A cell does not merely want to sense the presence of nutrients but wants to move toward the highest concentration. It does this by an amazing process that is sort of a primitive memory, where it essentially asks if its receptors are binding more or less of the compound than they were a moment ago (a moment ago being about 200 milliseconds.). If they are binding more, the cell keeps swimming, but if they are binding less, it tumbles and then swims in a random direction. This tumbling is the direct result of E. coli's ability to reverse the rotation of its flagella.
Phototaxis is somewhat different, but again the cells move toward optimal levels of specific wavelengths of light. Rhodobacter sphaeroides (a photosynthetic microbe) performs phototaxis by a mechanism analogous to chemotaxis in E. coli, but there are important differences. R. sphaeroides contains only one polar flagellum, which rotates during a run toward light. If, during a run, light conditions worsen, the flagella stop rotating (instead of reversing its rotation). The microbe depends upon Brownian movement in the environment to turn it in a new direction.
Aerotaxis refers to the ability of some bacteria to respond to the presence of oxygen. It is mechanistically generally similar to what we previously described and depends on dissolved oxygen levels in the environment. Whether a bacterium swims toward it or away depends on the type of metabolism that it has.
Some bacteria move by gliding motility
Flagella are not the only means bacteria have developed for moving about the environment. A large collection of phylogenetically diverse bacteria have developed gliding motility, an energy-requiring process by which bacteria move smoothly over a solid surface. Gliding microbes tend to be predominately Gram-negative, but there are examples of Gram-positive bacteria. This type of motility is not as well understood as flagellar propulsion, and there appear to be several different mechanisms employed to accomplish it.
In one mechanism, the microbes use pili (type iv pili specifically) that are extended away from the cell and stick to the surrounding surface. The microbe then pulls itself toward the tethered end by retracting the pilus back inside the cell. By repeating this process, the cell drags itself along a surface. This type of motility is observed in a number of microorganisms including Pseudomonas aeruginosa, Neisseria gonorrhoeae, and Myxococcus Xanthus. Movement in this manner requires energy in the form of ATP.
There are also gliding bacteria that use other mechanisms. Some filamentous cyanobacteria seem to extrude an extracellular polysaccharide through small pores on the surface of the organism. Figure 3.31 depicts this type of gliding motility. Researchers think that the polysaccharide exiting from the cell propels the microbe along the surface by some means.
Figure 3.31. Gliding motility in Myxococcus xanthus.. Vegetative cells of a Myxococcus xanthus moving across an agar surface using gliding motility.
A third groups of microbes (Cytophaga and Flavobacterium) seem to use yet another mechanism that is dependent upon proton motive force. Observation of these microbes in the presence of tiny latex beads shows the beads moving along the microbe's surface directionally. The bead moves from one pole of the rod-shaped bacteria to the other, sometimes reversing direction. One model to explain this behavior is the concerted movement of cytoplasmic membrane proteins that are coupled to outer membrane proteins. These membrane proteins might form a kind of conveyor belt. The outer membrane proteins in this mechanism contact the environmental surface, and their movement propels the microbe forward.
Interestingly, Myxococcus xanthus uses several of these mechanisms in its motility. M. xanthus is a social predator. It glides around in large groups of cells secreting toxins and degradative enzymes that kill other microbes. The leftovers of these dead microbes then provide nutrients for the marauding Myxococcus. Investigations by Dale Kaiser and others have revealed that this microbe has two types of gliding motility, social and adventurous. Social motility occurs in groups, while adventurous motility involves single cells. It turns out that social motility is dependent upon pili, as described above, while adventurous motility involves the extrusion of slime from pores in the microbe's cell wall.
Pili
Pili and fimbriae are structurally similar to flagella and are composed of one or a few proteins arranged in a helical fashion. Figure 3.32 shows pili isolated from Neisseria gonorrhea. Each protein subunit assembles on the growing structure at the tip, as is the case with flagella. There are many genes necessary for the successful construction of pili. Their products might perform functions such as moving the structural proteins across the membrane, methylating the structural proteins, or retracting the pilus. The same is generally true for fimbriae.
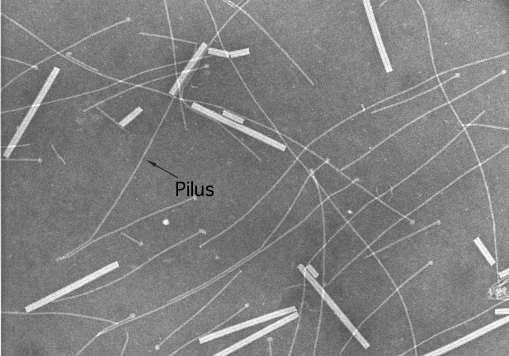
Figure 3.32. Pili and fimbriae. The pili shown in this micrograph are those of Neisseria gonorrhea with Tobacco Mosaic virus (the thicker structures) added as a size reference. The width of the Tobacco Mosaic virus is 0.018 µm. (Source: Katrina Forest, University of Wisconsin-Madison)
Fimbriae are found on many bacteria, are shorter and straighter than flagella, and more numerous. Not all bacteria synthesize them. Fimbriae do not function in motility but seem to be important in attachment to surfaces. Some microbes attach to hosts by fimbriae, and successful colonization of many surfaces depends on the ability to make fimbriae. Swarming microbes such as Myxococcususe them to sense the presence of similar microbes, which helps keep their "hunting packs" together.
Pili are longer than fimbriae, and there are only a few per cell. There are two basic functions for pili: gene transfer and attachment to surfaces. In gene transfer, the pilus captures a recipient bacterium, and DNA will be transferred from the donor to the recipient microbe. Conjugation, as this transfer process is known, is one explanation for the rapid spread of drug resistance in many different species of bacteria and is covered in depth in the Chapter on Genetics and Genomics.
Pili are also important for the attachment of many types of microorganisms to surfaces. For example, Neisseria gonorrhoeae, the causative agent of gonorrhea, has a special pilus that helps it adhere to the urogenital tract of its host. The microbe is much more virulent when able to synthesize pili.
Bacterial cells are often covered in glycocalyx
The general term for any network of polysaccharides or protein-containing material extending outside the cell is the glycocalyx. Many bacteria produce such a coating on the outside of their cell, and they come in two types: capsules and slime layers. The difference between the two is somewhat arbitrary. A capsule is closely associated with cells and does not wash off easily, while a slime layer is more diffuse and easily washes away. Figure 3.33 shows the capsule surrounding Klebsiella planticola.
Figure 3.33. Capsule surrounding cells of Klebsiella planticola.. The capsule (made of polysaccharide) in the figure is colorless and about the diameter of the cell. The background is darker.
There are many different types of proteins, polysaccharides, polyalcohols, and amino sugars in glycocalyx, and the exact makeup is species-specific. The structure can be thick or thin, rigid or flexible. Both Bacteria and Archaea are known to make capsules that will surround the cell.
There are several functions attributed to glycocalyx, one of which is to help cells attach to their target's environment. Streptococcus mutans produces a slime layer in the presence of sucrose. This forms a surface that allows other bacteria to aggregate on tooth surfaces and results in dental plaque. Vibrio cholerae, the cause of cholera, also produces a glycocalyx that helps it attach to the intestinal villi of the host. Glycocalyx can play other roles in pathogenesis as well. Bacteria that enter the body are always in danger of being attacked by phagocytes (host cells that protect you from invaders). Often the capsule makes it more difficult for phagocytes to attach to and engulf pathogens. As one example, Streptococcus pneumoniae, when encapsulated by a glycocalyx, can kill 90% of infected animals, while non-encapsulated forms cannot. In addition, capsules and slime layers are largely hydrophilic, so they can bind extra water in the environment and contribute to the protection of the cell from desiccation. Capsules and slime layers can also provide protection from the loss of nutrients by holding them within the layer. These extra layers coating the surface of the cell may also potentially mask viral receptors making it more difficult for viruses to attach. Many of these functions manifest themselves in biofilms. The glycocalyx is the glue that holds together whole communities of microorganisms. We will discuss biofilms in more detail in the chapter on Environmental Microbiology.
Key Takeaways
- Outside of the cell wall, bacteria contain structures that help them interact with their environment.
- For motility, microbes often use flagella, which can help them to move toward helpful environments and away from damaging environments.
- Other microbes can move using gliding motility, a method that depends upon pili or the excretion of slime.
- Pili can also be used for attachment to surfaces or as a method of exchanging genetic information.
- Fimbriae are shorter than pili, but can also serve at organs of attachment.
- Bacteria will also excrete a glycocalyx, a layer composed of protein or polysaccharide that helps the microbe stick to surfaces, and protect itself. Glycocalyx is important in the formation of biofilms.