Latest News
- Clues beginning to emerge on asymtomatic SARS-CoV-2 infection
- Back in November of 2020, during the first wave of the COVID-19 pandemic, I was teaching an in-person microbiology laboratory. One of my students had just been home to see his parents, and they all c…
- Read more
- Could there maybe be better uses of genetics and probiotics?
- Professor Meng Dong and his laboratory have created a probiotic that can metabolize alcohol quickly and maybe prevent some of the adverse effects of alcohol consumption. The scientists cloned a highl…
- Read more
- ChatGPT is not the end of essays in education
- The takeover of AI is upon us! AI can now take all our jobs, is the click-bait premise you hear from the news. While I cannot predict the future, I am dubious that AI will play such a dubious role in…
- Read more
- Fighting infections with infections
- Multi-drug-resistant bacterial infections are becoming more of an issue, with 1.2 million people dying of previously treatable bacterial infections. Scientists are frantically searching for new metho…
- Read more
- A tale of two colleges
- COVID-19 at the University of Wisconsin this fall has been pretty much a non-issue. While we are wearing masks, full in-person teaching is happening on campus. Bars, restaurants, and all other busine…
- Read more
16-4 T Lymphocytes Regulate the Immune System and Fight Intracellular Infections, Viral Infection and Cancer
( 32121 Reads)
|Learning Objectives
- Describe T cells and T cell receptors.
- Explain how T cells interact with antigen presenting cells.
- Describe how T cell mature in the thymus.
- Explain the role of major histocompatibility complex proteins, MHC I and MHC II in the immune response.
- Explain the mechanism of T cell activation.
- Describe the role of cytokines and chemokines.
T lymphocytes or T cells are the second arm of the immune system and are part of what is called cell-mediated immunity. Their roles are to regulate the immune system, help B lymphocytes in the production of antibodies and deal with threats to our body that involve our own cells (e.g., intracellular bacterial infections, viral infections and cancer). T cells recognize antigens in our bodies using receptors on their surface, sensibly called T cell receptors (TCR) as shown in Figure 16.6. These proteins are somewhat reminiscent of antibodies and are composed of two protein fragments, α and β. Each polypeptide chain can be broken into three domains. The carboxy-terminal domain serves as a hydrophobic anchor in the cellular membrane and also contains a disulfide bridge near the outside membrane surface that links the α and β polypeptides. The middle of each protein forms a constant region that shares homology across all TCRs and serves as scaffold for the variable domain projecting furthest into the surrounding environment. TCRs occur in association with CD3 (another protein on the surface of T cells) to form a TCR-CD3 complex. It is this complex that recognizes presented antigens and begins the T cell activation process.
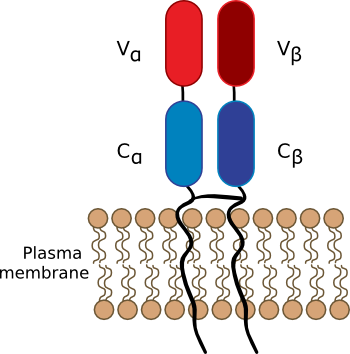
Figure 16.6. The T-Cell Receptor. The structure of the T cell receptor (TCR) is reminiscent of antibody structure. However, T cell receptors are found anchored in the membrane of T cells, while the majority of antibodies float free in various fluids of the body. (Left panel) a cartoon showing the constant and variable regions of the TCR. (Right panel) A molecular model of a TCR. (Source: L. Kjer-Nielsen, et al. 2003. Immunity 18:53–64)
The variable regions of these receptors react with antigens in a fashion that is reminiscent of antigen-antibody reactions. TCRs can respond to a large assortment of molecules and this diversity is achieved by a mechanism similar to that used to create antibody diversity as discussed previously. In contrast to antibodies, TCRs respond to antigens only when it is associated with a membrane protein of another host cell such as antigen-presenting cells (APC). These APCs wrap fragments of foreign molecules inside special proteins, called Major Histocompatibility Complex (MHC) molecules and then display them on their surface for presentation to T cells. In order to understand the mechanism of T cell activation, it is useful to first learn about these important protein complexes present on every animal cell.
Major Histocompatibility Complex Molecules Are the Signal Beacons of the Immune System
Almost all human tissues contain Major Histocompatibility Complex (MHC )molecules, which serve as signal beacons for the immune system. There are two types of MHC molecules. MHC I molecules are found on almost all types of body cells, whereas MHC II molecules are only found on macrophages, dendritic cells and B cells, which can serve as professional antigen-presenting cells. MHC I molecules are status monitors for all cells of the body, while MHC II molecules are important in presenting foreign antigens to other cells of the immune system.
The MHC I molecule consists of a α chain and a β2-microglobulin. One segment of the α chain is anchored to the membrane, with the rest of the molecule protruding outside the cell. Class II MHC molecules are also transmembrane proteins consisting of α and β chains. The two chains fold to give a structure somewhat similar to that of the MHC I molecule, except that both α and β chains are anchored in the membrane. Both MHC I and MHC II molecules contain a deep groove in the domain of the molecule farthest away from the membrane and therefore exposed to the environment. This groove is capable of binding short peptides or other antigens that are not part of the MHC molecule. The peptides in the groove are from various sources, as you will see below. The MHC I groove is closed off and can contain only short peptide sequences, while the MHC II molecule is open and can bind to peptides of longer lengths. Figure 16.7 depicts the structure of MHC proteins.
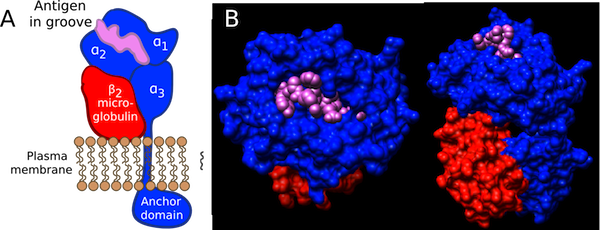
Figure 16.7. Structure of an MHC I Molecule. Major histocompatibility complex proteins are found on most cells of the body. They function to display proteins to T cells. MHC I molecules display proteins that originate in the cytoplasm of the displaying cell. A) A cartoon of the domains of an MHC I molecule. B) Top and side views showing the binding of an antigen (in this case, an influenza peptide) to mouse MHC I molecule. The anchor domain is not shown in the molecular model. (Source: R. Meijers, et al.. 2005. J. Molec. Biol. 345:1099-1110)
Each type of MHC molecule binds peptides from a different source, consistent with their different roles in the immune system. MHC I molecules bind to peptides that originate in the cytoplasm. These peptides were part of cytoplasmic proteins, but were digested by the cellular degradative machinery. This machinery is part of the normal function of a cell and serves to recycle old protein making it available for future use. Some of the short peptide fragments that result from this process are recognized by a specific transporter molecule and translocated from the cytoplasm into the endoplasmic reticulum. Here they are picked up by newly synthesized MHC I molecules and move to the cellular membrane. In this way, the MHC I molecules serve as flags indicating what type of protein processing is going on inside the cell. If the cell is functioning normally, the MHC I molecules contain inside their grooves short peptides derived from normal cell proteins. The immune system recognizes these proteins as self and presumes that all is normal with the cell. In contrast, if the cell is infected by an intracellular parasite, such as virus, some of the MHC I molecules are bound to contain peptide fragments from the virus. These molecules then interact with T cells of the immune system and initiate a response.
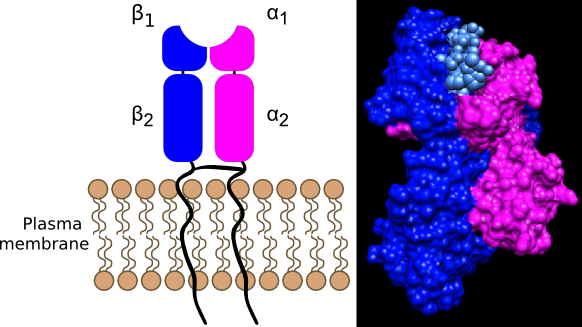
Figure 16.8. An MHC II molecule. MHC II molecules found only on antigen-presenting cells, display foreign antigens picked up in the environment. They contain an α and a β domain. The α1 and β1 domains anchor the protein to the membrane while the α2 and β2 domains form the groove where antigens bind. The MHC II groove is open allowing for the insertion of a larger fragment, 15 to 24 amino acids long. The left panel shows a cartoon of a MHC II molecule, while the right shows the human MHC II molecule binding an antigen (Source Li et. al (2010) J.Mol.Biol. 399: 596-603)
MHC II molecules are present on cells that are part of the immune system and serve as vessels to present foreign antigens to T cells. The structure of the MHC II molecule is shown in Figure 16.8. MHC II molecules bind to antigens that originate from an exogenous source, for example, the leftovers of a phagolysosome after a microorganism has been killed and degraded. Peptide fragments coming from the phagolysosome are combined with preformed MHC II molecules and move to the cell surface. The MHC II molecule, together with the antigen, is then recognized by T cells of the immune system.
There Are Several Types of T Cells, but All Mature in the Thymus
The response T cells have to the presentation of antigens by the MHC molecules on other cells depends upon the type of T cell. Two major subpopulations of T cells differ from one another by the type of surface protein that they display. Mature T cells contain either the CD4 protein or the CD8 protein on their surface. Cells that contain CD4 are further subdivided into four subsets: TH0 cells; T helper 1 cells (TH1); T helper 2 cells (TH2); and pathogenic T cells (T17);. When TH0 cells encounter an antigen they react with, they differentiate into TH1, TH2 or T17 cells, depending the cytokines in the environment, which in turn depends upon the nature of the antigen. TH1 cells participate in cell-mediated immunity and are responsible for recruiting and regulating nonspecific immune cells such as phagocytes. They also help to regulate the steps of inflammation and thus are called T inflammatory cells. TH2 cells serve to activate B cells to differentiate and produce antibody. In most cases, only small amounts of antibody are produced by B cells on their own and TH2 cells are necessary to get full activation of the B cells and large amounts of antibody production. Not much is known about T17 cells, but it is know that their activation increases the severity of some autoimmune diseases (inflammatory bowel disease, multiple sclerosis and rheumatoid arthritis)
The second group of T cells, those with CD8 on their surface, are called T cytotoxic (Tc) cells, also known as cytotoxic T lymphocytes. Mature Tc cells cannot kill target cells until they are activated. This is a two step process.
- The Tc cell has to recognize an antigen presented by a MHC I molecule on a host cell
- The Tc cell has to then be exposed to cytokines (especially IL-2 and IFN-gamma) produced by TH1 cells.
Subsequent presentation of a recognized antigen by a host cell to a Tc cell causes it to attack and kill that cell. The killing involves two mechanisms.
- Tc cells secrete Fas and tumor necrosis factor (TNF) ligands on their surfaces that bind to Fas and TNF receptors on the surface of the target cell. This precipitates a signaling pathway that leads to apoptosis (cell death).
- Fully differentiated Tc cells have granules inside them containing perforin and granzymes. The Tc cell makes contact with the target cell and then releases the perforin. This polymerizes and forms pores in the target cell membrane. The granzymes then release from the Tc cell and migrate through the perforin pores, into the target cell. Once there, the granzymes (serine proteases) activate nucleases and caspases in the target cell resulting in apoptosis.
Tc cells can be thought of as sentries that follow prescribed routes, patrolling our bodies looking for cells displaying odd antigens in their MHC molecules. They also respond to cells that raise the alarm, sent out in the form of an inflammatory response. When found, Tc cells destroy these cells.
T Cell Maturation and Tolerance
T cells originate from bone marrow stem cells in an immature form. Immature T cells can collectively attack a wide range of antigens, including antigens on our own cells. These cells must be selected to only react against non-self antigens and this education takes place in the thymus as shown in Figure 16.9.
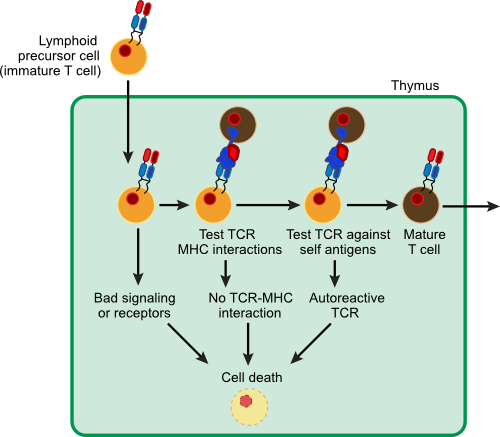
Figure 16.9. T Cell Maturation. Immature T cells move to the thymus for their maturation. The maturation process occurs in two steps. There is a positive selection to ensure that the T cell can interact with MHC molecules correctly. Then there is a negative selection to eliminate those T cells that interact with MHC molecules containing self antigens. T cells that survive this process (only 10%) mature and migrate out of the thymus.
At this point, the education of the T cells begins. Remarkably, immature T cells are initially programmed to die and only through intervention by the thymus do they survive. During their time in the thymus, T cells are exposed to various self-antigens. If they react to the antigen, the T cell is encouraged to carry out its cellular program and die. In fact, the thymus is something of a graveyard for T cells with only about 10% surviving the treatment (some education system!). It may seem more logical to set up the system so that the thymus kills bad T cells, but if you think about it, preprogrammed death is actually safer. If a damaging immature T cell somehow escapes education, it will still die since an educating cell did not tell it to live. In this fashion, the body averts many potential autoimmune reactions. Those T cells that do survive are assumed to be protective against non-self antigens and are released into the bloodstream. These mature T cells then roam throughout the body to fulfill their functions.
Note that B cells undergo a similar type of culling process as T cells to remove self-reactive antibodies, but this process takes place in the bone marrow and not in the thymus. Since the process is very similar to that described for T cells, we will not cover it in detail. Briefly, developing B cells are presented with self-antigens. Reaction between the B cell receptor (remember this is IgD or IgM on the surface of the B cell) and the antigen causes the B cell to die. In this way, the body only produces antibodies to foreign antigens.
The above mechanism is one way the body creates tolerance to self-antigens, but the process is not foolproof. The thymus does not have all the self-antigens of the body available to it and rarely a T cell matures that can potentially react with various parts of the body. In these cases, other mechanisms cope with the presence of a self-reactive T cell, including clonal anergy, active suppression and sequestering.
Clonal anergy is a mechanism where T cells are made unresponsive to antigens, but are not killed. This situation is brought about by T cells reacting with antigen presented on MHC I molecules, but not receiving other signals from the immune system to multiply. Activation of T cells requires presentation of the antigen in the context of a MHC II molecule and a second receptor only found on antigen-presenting cells. Since these are self-antigens, they are not presented to the T cells by an antigen-presenting cell and are only detected on MHC I molecules. The absence of this second signal causes the T cell to shut down and become incapable of releasing IL-2 even during subsequent exposure to the antigen. Since cells expressing MHC I molecules, but not the second signal, are present throughout the body, many self-antigens are readily encountered by a reactive T cell and it is quickly made unresponsive.
Certain T cells, called T suppressor cells, are necessary for turning down the immune system after the source of an antigen has been dealt with. Once a viral or bacterial infection is eliminated, the immune system must be suppressed to prevent further tissue damage. Remember that many of the immune reactions are themselves very damaging to tissue and it is imperative to stop these reactions as soon as they have fulfilled their purpose. T suppressor cells also have a role in protection from self-reactive antigens by mechanisms termed active suppression. It is known that normally healthy humans contain T cells that can potentially cause autoimmune disease and these cells are prevented from causing harm by the action of T suppressor cells. It has also been suggested that T suppressor cells can be induced by reaction with specific antigen, in this case a self-antigen, so that the suppression response matches any unwanted autoimmune reaction of other cells. The mechanism of how this whole process works is still not clear.
A final mechanism to deal with potential autoimmunity is the sequestering (hiding) of potentially stimulatory antigens. Some sites of the body are considered off-limits to the immune system either because the immune system cannot reach the site or for some reason, immune cells cannot detect the antigen due to poor presentation. The classic example is the cornea of the eye. There are no blood vessels that run through this structure, so immune cells have no access to its antigens and therefore, no immune response can be raised. Thus, it is possible to transplant the cornea without fearing rejection by the recipient's immune system. In other cases, certain antigens are hidden away inside cells so that the immune system does not have access to them. When all of these systems fail, it can result in the manifestations of autoimmune disease and we will discuss these later in the chapter.
T Cell Activation Occurs when an Antigen Is Presented to a T Cell and Reacts with Its T Cell Receptor
T cell activation, Figure 16.10, follows a similar pathway to that seen for B cell activation. The best-known activation pathway is that of IL-2 synthesis by TH1 cells and we will go through it in some detail to give you an idea of the molecular mechanisms involved in the process. Similar pathways are present for the activation of other cytokines.
Figure 16.10. Activation of Il-2 Expression in a Helper T Cell. If the TCR recognizes the antigen, a complex is formed between the MHC II molecule, the TCR and the CD4 protein. Successful docking activates a tyrosine kinase in the cytoplasm and this begins a series of cellular reactions. Tyrosine kinase phosphorylates a tyrosine residue on phospholipase Cg1 causing it to become active and split phosphatidyl inositol bisphosphate into inositol trisphosphate and diacylglycerol. Inositol trisphosphate causes a calcium channel to open in the cell membrane resulting in an influx of calcium. The increased calcium levels cause the activation of calmodulin, calcineurin and eventually, nuclear factor of activated T-helper cells (NF-AT). Diacylglycerol activates protein kinase C, which moves into the nucleus and catalyzes the formation of the AP-1 complex from the JUN and POS proteins. NF-AT and AP-1 then form a complex (a transcription factor) that can recognize and bind to a DNA sequence near the IL-2 gene. A second signal is also necessary to activate the T cell. Antigen-presenting cells contain the protein B7-2 on their membranes, which reacts with the CD28 receptor on the T cell. Binding of B7-2 to CD28 activates a separate tyrosine kinase that phosphorylates CD28RC. This protein binds to a site on the IL-2 gene enhancing transcription and also stabilizes IL-2 mRNA once it is formed, increasing its functional concentration. This secondary signal greatly magnifies IL-2 mRNA expression. IL-2 mRNA then moves out of the nucleus for translation at the ribosome.
An antigen-presenting cell that contains a foreign antigen in its MHC II molecule first comes into cell-to-cell contact with a T helper cell and its TCR. Several signals are necessary to activate a T cell. Close contact between the helper T cell and the antigen-presenting cell is required. The first signal is the complex between the MHC II molecule and the TCR, causing the eventual formation of a DNA binding protein (NF-AT/AP-1) that enhances IL-2 transcription. The second signal is between B7-2, another membrane protein on the antigen-presenting cell and the CD28 receptor. This creates a separate signal that results in the formation of a second effector protein, CD28. This then binds to the DNA near the IL-2 gene and also stabilizes the IL-2 mRNA. Both signals, that from the MHC II molecule and the B7-2 protein, are necessary to fully activate the T helper cell.
Another important group of signal proteins in immunity are the chemokines. Chemokines are a set of small molecular weight proteins (8-12 kDa) that serve as chemoattractants to phagocytes and T cells. Damage to cellular tissue from bacterial invasion, viral infection or physical injury causes the secretion of chemokines. Since many parts of the body are susceptible to this type of damage, it is not surprising that a large assortment of cell types, including lymphocytes, are capable of synthesizing chemokines. Their production attracts T cells and phagocytes to the area of damage stimulating an inflammatory and immune response.
A key feature of the adaptive immune response is its memory. After encountering a pathogen and successfully fighting it off, T- and B-memory cells are distributed throughout the body. Upon subsequent challenge by the same pathogen having similar antigens, a much stronger and more rapid response occurs. The enhanced response to a subsequent infection is called a secondary response. It stops the pathogen before it can cause disease. The secondary response is the reason immunity to a pathogen develops. Figure 16.11 shows antibody synthesis during a primary and secondary response.
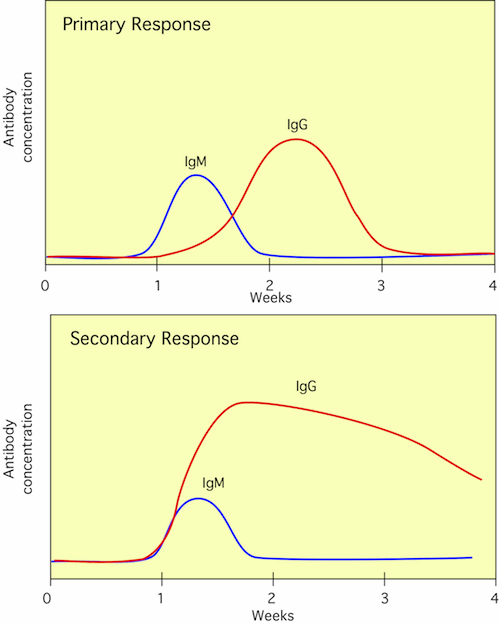
Figure 16.11. A Graph of Antibody Responses IgM and IgG Titers with 1° and 2° Responses to Antigen.. In a first response to antigen (primary response), the antibodies appear more slowly, at a lower concentration, and contain a higher proportion of IgM. During a second exposure to antigen (secondary response) the antibody appears more quickly, reaches a higher concentration and contains more Ig.
Natural Killer Cell Activation Involves Interferon or Il-2, and IgG
We now again return to natural killer cells. Natural killer cells cannot be induced as cytotoxic T cells are but they can be activated by the mechanism of Antibody-Dependent Cell-Mediated Cytotoxicity, also know as ADCC. The mechanism of natural killer attack involves several separate activation steps. Interferon from virally infected cells or IL-2 from T cells must first activate natural killer cells. The second activation step occurs when the natural killer cells encounters a target cell coated with IgG and docks with its Fc region. Having several activation steps prevents the natural killer cell from mistakenly attacking normal cells. Docking then causes changes in the location of cytoplasmic granules and Golgi apparatus within the natural killer cell to a location close to the edge of the cytoplasmic membrane and near the target cell. At this point, the natural killer cell is activated and ready to attack its target. In a mechanisms very similar to that for Tc cells, a pore-forming protein, perforin 1 is inserted into the membrane of the target and the natural killer cell then exports lysosomal secretions into the target cell. The pore made in the membrane together with the attack of the lysosomal contents eventually causes the target cell to lyse and die. The natural killer cell then prepares its cytoplasmic components for the next victim.
Key Takeaways
- T cells respond to antigens using a T cell receptor that is similar in structure to antibodies. Antigen presenting cells present antigen to T cells inside a major histocompatibility complex protein. MHC molecules are membrane-bound proteins: MHC I molecules are found on almost all tissues of the body, while MHC II molecules are found only on antigen-presenting cells.
- MHC molecules possess a deep groove that is capable of holding a short peptide. MHC I molecules process proteins present inside the cell and present them on their surface. MHC II molecules present antigens taken from the phagosome digestion, most often foreign cells, and present them to the immune system. The immune system monitors the proteins present on MHC I molecules and activates when a foreign protein, from an intracellular parasite, is detected. This normally results in the destruction of the cell.
- Mature T cells are classified by their role the immune system. T helper cells help regulate the immune system. Cytotoxic T cells attack cells that have been infected or are displaying odd antigens. In the thymus, T cells are exposed to various self-antigens. If they react, the T cell is allowed to die, if they do not, then the T cell survives. Not all host-reactive T cells are eliminated by their education in the thymus, but these self-reactive T cells are kept under control by clonal anergy, active suppression and sequestering.
- T cell activation occurs when an antigen the T cell reacts to is presented to it in the context of a MHC II molecule. As one example in TH1 cells, binding of T cell receptor to a MHC II molecule causes the formation of proteins that activate DNA transcription of the IL2 gene. IL2 is one example of a cytokine.
- Natural killer cells are activated when a chemokine (either interferon or IL-2) is present and IgG is recognized on the surface of a cell. This causes the natural killer cell to attack, forming a pore in the target cell and pumping lysosomal contents into it causing its death.
Quickcheck 16-4
Warning, you must be logged in to be able to have your exam graded. Answer the questions below and if you are a registered user of the site you will see a Grade Exam button. Click it to have your exam graded.