Latest News
- Clues beginning to emerge on asymtomatic SARS-CoV-2 infection
- Back in November of 2020, during the first wave of the COVID-19 pandemic, I was teaching an in-person microbiology laboratory. One of my students had just been home to see his parents, and they all c…
- Read more
- Could there maybe be better uses of genetics and probiotics?
- Professor Meng Dong and his laboratory have created a probiotic that can metabolize alcohol quickly and maybe prevent some of the adverse effects of alcohol consumption. The scientists cloned a highl…
- Read more
- ChatGPT is not the end of essays in education
- The takeover of AI is upon us! AI can now take all our jobs, is the click-bait premise you hear from the news. While I cannot predict the future, I am dubious that AI will play such a dubious role in…
- Read more
- Fighting infections with infections
- Multi-drug-resistant bacterial infections are becoming more of an issue, with 1.2 million people dying of previously treatable bacterial infections. Scientists are frantically searching for new metho…
- Read more
- A tale of two colleges
- COVID-19 at the University of Wisconsin this fall has been pretty much a non-issue. While we are wearing masks, full in-person teaching is happening on campus. Bars, restaurants, and all other busine…
- Read more
( 61127 Reads)
|Learning Objectives
After reading this section, students will...
- Be able to describe the function and location of the periplasm.
- Be able to describe the function of the cell wall and the two common cell wall structures.
The periplasm is between the cytoplasmic and outer membranes in Gram-negative bacteria
The periplasm is found in Gram-negative bacteria and is the space in between the cytoplasmic and outer membranes. (Some feel a periplasm-like compartment is also present in Gram-positive bacteria between the cytoplasmic membrane and the peptidoglycan.) The periplasm is filled with water and proteins and is therefore somewhat reminiscent of the cytoplasm. However, pools of small molecules in the periplasm are not like those in the cytoplasm because the membrane prevents the free exchange between these two compartments. Also, the proteins found in the periplasm are distinct from those in the cytoplasm. They are specifically guided to this site during translation through specific <a class="glossary">signal sequences typically near their N-termini. Table 3.2 lists some examples of these proteins.
The peptidoglycan shell that provides the strength to prokaryotic membranes is also found in the periplasmic space of Gram-negative bacteria. In contrast, in Gram-positive bacteria, peptidoglycan provides the outside border to the periplasm.
Table 3.2 Different types of periplasmic enzymes and their role in the cell
| Enzyme Type | Examples | Function |
| Hydrolytic enzymes | phophatases | Degrading phosphate-containing compounds. |
| proteases | Degrading proteins and peptides. | |
| endonucleases | Degrading nucleic acids. | |
| Binding proteins | sugars, amino acids, norganic ions, vitamins | Binding substrates and docking with transport protein in membrane. |
| Chemoreceptors | Chemotaxis, ermination | Sensing the environment and changing cell behavior in response. |
| Detoxifying enzymes | β-lactamase | Degrading penicillin and related compounds before they get into the cell. |
Periplasmic enzymes have several main functions, detecting nutrients in the environment, degradation of polymers, and protection from harmful compounds.
The cell wall surrounds and holds in the microbe
This section will restrict itself to the bacterial cell wall. We will compare this to archaeal cell walls at the end of the chapter. The cell wall is essential to the survival of most microorganisms. Many microbes live in relatively dilute environments, and the wall's most important function is to prevent the cell from bursting due to osmotic stress. The cell wall also determines the shape of the cell. Any cell that has lost its cell wall, either artificially or naturally, becomes roughly spherical and lyses due to osmotic pressure unless placed in certain concentrated solutions. Finally, the cell wall helps to support any structure that penetrates from the cell out into the environment.
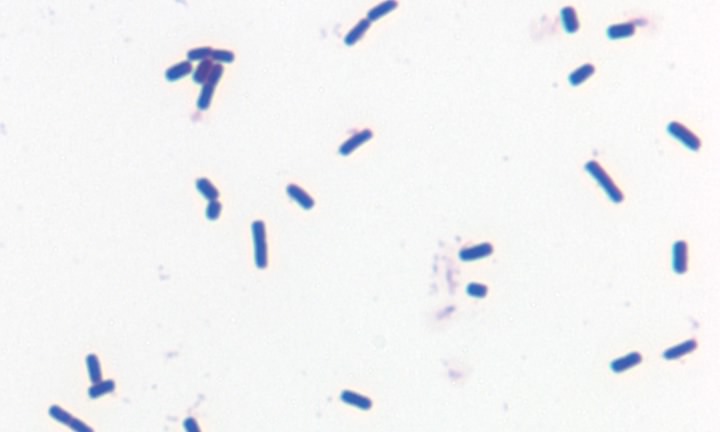
Figure 3.19. A gram-positive bacterium. Gram stain of the gram-positive bacterium Bacillus cereus
The structure and synthesis of prokaryotic cell walls is unique, and many compounds found in the bacterial cell wall are found nowhere else in nature. It is true that plants also make cell walls, but they are chemically and structurally different. Investigations have revealed two basic types of bacterial cell wall structures: Gram-positive and Gram-negative. These two classes of bacterial cells look very different following staining with the Gram stain, and this has been a standard test for identification of bacterial species. Figures 2-19 and 2-20 show Gram stains of Gram-positive and Gram-negative bacteria, respectively.
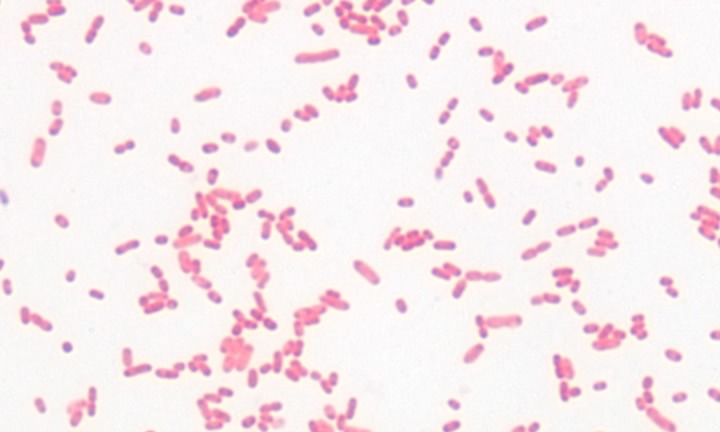
Figure 3.20. A gram-negative bacterium. A Gram stain of the gram-negative bacterium Serratia marcescens
When Hans Christian Gram developed the Gram stain in 1884, the molecular basis of the stain was unknown. Bacteriologists understood very little about bacteria in general. He just empirically determined that when bacterial smears were run through a four-step staining procedure using two different dyes, some cells retained the first dye and stained purple, while others only retained the second dye and stained pink. Years later, bacteriologists discovered that this differential reaction relates to the cell wall, as shown in Figure 3.21.
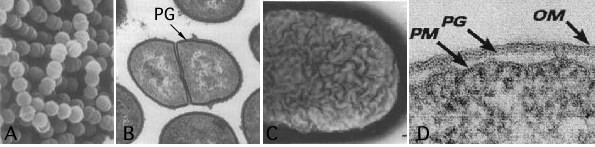
Figure 3.21. A comparison of the ultrastructure of gram-positive and gram-negative cells. The different Gram reactions occur because of structural differences between the bacterial cell walls. Gram-positive cells (Group B streptococci) appear smooth in a scanning electron micrograph (A) and are composed of a single layer of peptidoglycan (B). Gram-negative cells (E. coli) have an undulating surface and have three layers (C and D). (Sources: S. H. Pincus, et al.1992. J. Bacteriol 174:3739-3749 [panels A and B]; M. E. Bayer and C. C. Remsen. 1970. J. Bacteriol. 101:304-313 [panel C]; T. J. Beveridge. 1999. J. Bacteriol. 181:4725-4733 [panel D])
As shown in Figure 3.21, the Gram-negative cell has an additional layer, and the outside of the cell appears convoluted when compared to the Gram-positive cell. The Gram-positive wall is much thicker than is the Gram-negative wall, and its external appearance is smoother. Gram-positive and Gram-negative cells do share one thing in common that is unique to bacteria - peptidoglycan. We will talk about the structure of this and then examine the various structures found in each cell wall type.
Peptidoglycan is a thick rigid layer composed of an overlapping lattice of two sugars, N-acetyl glucosamine (NAG) and N-acetyl muramic acid (NAM), that are cross-linked by amino acid bridges, as shown in Figure 3.22. The exact molecular makeup of these cross-bridges is species-specific. NAM is only found in the cell walls of bacteria and nowhere else. Attached to NAM is a side chain generally composed of four amino acids. In the best-studied bacterial cell walls (E. coli), the cross-bridge is most commonly composed of L-alanine, D-alanine, D-glutamic acid, and diaminopimelic acid (DPA).
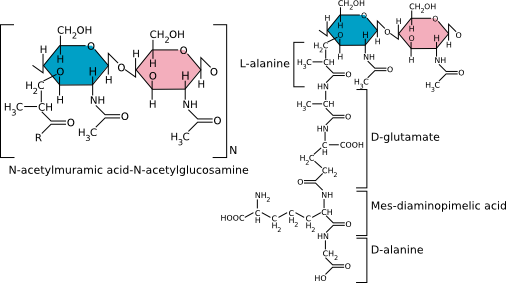
Figure 3.22. The chemical structure of peptidoglycan. The generalized peptidoglycan monomer showing the two sugars that make up the backbone. The R group consists of four amino acids, with the best-studied cell walls containing L-alanine, D-alanine, D-glutamic acid and diaminopimelic acid.
Note that peptidoglycan contains D-amino acids, which are different than the L-amino acids found in proteins. D-amino acids have the identical composition as L-amino acids but are their mirror images. The use of D-amino acids is unusual in biology, and bacteria have enzymes called racemases to convert between D and L forms specifically for this use.
The NAM, NAG, and amino acid side chain form a single peptidoglycan unit that can link with other units via covalent bonds to form a repeating polymer. Covalent bonds between cross-bridges further strengthen the polymer, and the degree of cross-linking determines the degree of rigidity. Figure 3.23 shows an artist's rendering of what the structure might look like. In E. coli, the penultimate D-alanine of one unit links to DPA of the next cross-bridge. In some Gram-positive microbes, there is a peptide composed of various amino acids that link the cross-bridges. For example, in Staphylococcus aureus strains, five glycines make up the linker between peptidoglycan monomers. The sequence of these linkers varies considerably between species. The completed peptidoglycan layer forms a strong mesh analogous to a chain-link fence. The complete cell wall contains one or more layers of peptidoglycan, one atop the other, providing much of the strength of the cell wall.
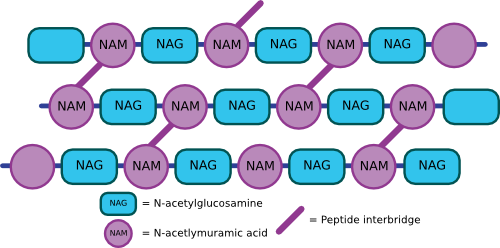
Figure 3.23. A cartoon of the peptiodglycan mesh. The peptidoglycan polymers then crosslink with other peptidoglycan chains to form a complex mesh that wraps the cell in a structure a kin to chicken wire.
While both Gram-negative and Gram-positive bacteria have peptidoglycan, their physical arrangement in the cell wall differs. In Gram-positive cells, the peptidoglycan is a heavily cross-linked woven structure that encircles the cell in many layers. It is very thick, with peptidoglycan accounting for 50% of the cell weight and 90% of the weight of the cell wall. Electron micrographs show the peptidoglycan to be 20-80 nm thick. In Gram-negative bacteria, the peptidoglycan is much thinner, with only 15-20% of the cell wall being peptidoglycan. In both cases, peptidoglycan is not a barrier to solutes, as the openings in the mesh are large enough for most molecules, including proteins, to pass through. Figure 3.24 shows a depiction of the Gram-positive cell wall.
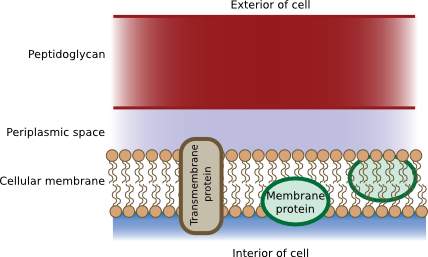
Figure 3.24. The Gram-positive cell wall. The cell wall is made mostly of peptidoglycan, interspersed with teichoic acid which knits the different layers together. The amount of crosslinking is higher and the wall is thicker than in gram-negative cell walls.
The Gram-positive cell wall
Gram-positive cells consist almost entirely of peptidoglycan, but teichoic acid is an important structure found in Gram-positive cell walls. It is a phosphodiester polymer of glycerol or ribitol joined by phosphate groups. Amino acids such as D-alanine are attached. Teichoic acid is covalently linked to muramic acid and stitches various layers of the peptidoglycan mesh together. Teichoic acid stabilizes the cell wall and makes it stronger. Figures 3.25 shows the chemical formula of teichoic acid.
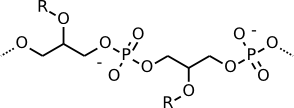
Figure 3.25. Teichoic acid. Teichoic acid is a long, thin molecule that weaves through the peptidoglycan.
Gram-negative cell structure
Gram-negative cell walls have a more complicated structure than that of Gram-positive organisms. Outside the cytoplasmic membrane is the periplasm, which contains the thin layer of peptidoglycan. The peptidoglycan in Gram-negative cells contains less cross-linking than in Gram-positive cells with no peptide linker. Covalently bound to the peptidoglycan is Braun's lipoprotein, which has a hydrophobic anchor that helps to bind the peptidoglycan to the outer membrane. Figure 3.26 shows the arrangement of the Gram-negative cell wall.
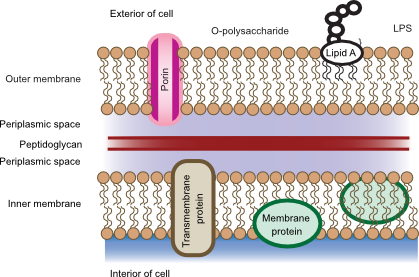
Figure 3.26. The Gram-negative cell wall. The cell wall in Gram-negative bacteria contains much less peptidoglycan and is surrounded by an outer membrane. There is much less crosslinking between the peptidoglycan. LPS is also present in the outer membrane and penetrates into the surrounding environment.
The outer membrane
The outer membrane of Gram-negative bacteria is another lipid bilayer similar to the cytoplasmic membrane, and contains lipids, proteins, and also lipopolysaccharides(LPS). It is a barrier to proteins and prevents enzymes secreted into the periplasm from floating away. The membrane has distinctive sides, with the side that faces the outside containing all the LPS. LPS is composed of two parts: Lipid A and the polysaccharide chain that reaches out into the environment. Lipid A is a derivative of two NAG units with up to 7 hydrophobic fatty acids connected to it that anchor the LPS in the membrane, as shown in Figure 3.27. Attached to Lipid A is a conserved core polysaccharide that contains KDO, heptose, glucose, and glucosamine sugars. The rest of the polysaccharide consists of repeating sugar units, and this is called the O-antigen. The O-antigen varies among bacterial species and even among various isolates of the same species. Many bacterial pathogens vary the makeup of the O-antigen to avoid recognition by the host's immune system.
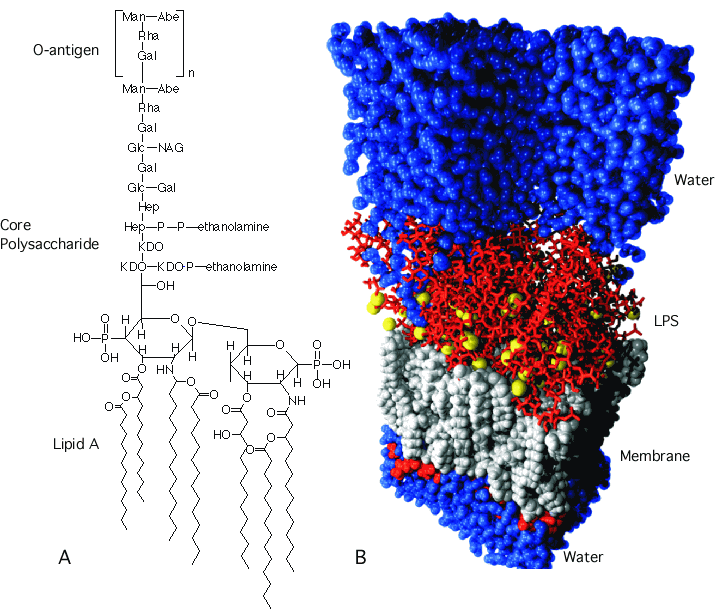
Figure 3.27. The structure of LPS. LPS is composed of three sections: the lipid A region, a conserved core polysaccharide, and a highly variable O-polysaccharide. (A) The chemical structure of LPS. (B) A molecular model of the outer membrane from Pseudomonas aeruginosa. Source (T. P. Straatsma, Pacific Northwest National Laboratory)
LPS confers a negative charge and also repels hydrophobic compounds, including certain drugs and disinfectants that would otherwise kill the cell. Some Gram-negative species live in mammals' gut, and LPS repels fat-solubilizing molecules such as bile that the gall bladder secretes. This repulsion enables these bacteria to survive in this environment. The O-antigen and other molecules on the outer membrane are targeted by certain viruses that infect bacteria as a means to identify the correct hosts for infection.
LPS is medically important because when LPS is released from bacterial cells it is toxic to mammals and is therefore called endotoxin. It creates a wide spectrum of physiological reactions including the induction of a fever (endotoxins are said to be pyrogenic), changes in white blood cell counts, leakage from blood vessels, tumor necrosis and lowered blood pressure leading to vascular collapse and eventually shock. At high enough concentrations the LPS endotoxin is lethal.
There are fewer total proteins and fewer unique types of proteins in the outer membrane than in the cytoplasmic membrane. Porins are essential components because of their role in the permeability of the outer membrane to small molecules. Porins form pores in the outer membrane large enough to allow the passage of most small hydrophilic molecules. Figure 3.28 shows the structure of a porin at the molecular scale. All known porins have a similar structure, with the protein containing a central channel that allows the passage of molecules. This common structure allows the migration of these molecules into the periplasmic space for possible transport across the cytoplasmic membrane. Some porins in the outer membrane are general, doing simple discrimination on size and charge, but having little substrate specificity. Examples include OmpF, which is selective for positively charged molecules, and PhoE, which is permeable to negatively charged molecules. Other porins are more specific. The best-studied is LamB, which recognizes the sugar polymer maltooligosaccharide and transports it through the outer membrane. Very large or hydrophobic molecules cannot penetrate the outer membrane, so the outer membrane serves as a permeability barrier to at least some molecules.
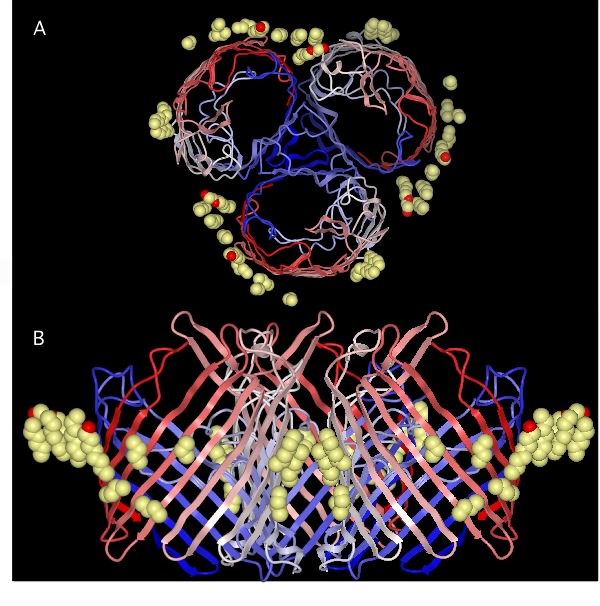
Figure 3.28. The molecular structure of a porin. The view in (A) is from the outside of the cell looking at the membrane surface. The view in (B) is the perspective from the side (i.e. from the membrane). The porin has three protein subunits and the actual pore is the central triangular area in the top panel formed by the three subunits.
Other types of outer membrane proteins are involved in various functions. OmpA in E. coli seems to connect the outer membrane to the peptidoglycan. Some pathogens contain outer membrane proteins that help them neutralize host defenses. Finally, all Gram-negative bacteria contain high molecular weight proteins involved in the uptake of large substrates such as iron-complexes and vitamin B12.
The differences between Gram-positive and Gram-negative bacterial cell walls greatly influence the success of the microbes in their environments. The thick cell wall of Gram-positive cells allows them to do better in dry conditions because it reduces water loss. The outer membrane and its LPS help Gram-negative cells excel in the intestines and other host environments. Table 3.3 summarizes the difference between Gram-negative and Gram-positive cell walls.
Table 3.3 Properties of cell walls
| Property | Gram-positive | Gram-negative |
| Thickness of wall | 20-80 nm | 10 nm |
| Number of layers in wall | 1 | 2 |
| Peptidoglycan content | >50% | 10-20% |
| Teichoic acid in wall | + | - |
| Lipid and lipoprotein content | 0-3% | 58% |
| Protein content | 0% | 9% |
| Lipopolysaccharide | 0 | 13% |
| Sensitive to penicillin | Yes | Less sensitive |
| Digested by lysozyme | Yes | Weakly |
A summary of the differences between Gram-positive and Gram-negative cell walls.
Some bacteria lack cell walls
For most bacterial cells, the cell wall is critical to cell survival, yet some bacteria do not have cell walls. Mycoplasma species are widespread examples, and some can be intracellular pathogens that grow inside their hosts. Cell walls are unnecessary here because the cells only live in the controlled osmotic environment of other cells. They could likely form a cell wall at some point in the past, but as their lifestyle became one of existence inside other cells, they lost the ability to form walls. Consistent with this very limited lifestyle within other cells, these microbes also have very small genomes. They do not need the genes for all sorts of biosynthetic enzymes, as they can steal the final components of these pathways from the host. Similarly, since their intracellular environment is completely predictable, they can also do without genes encoding many different pathways for various carbon, nitrogen, and energy sources. Because of the absence of cell walls, Mycoplasma have a spherical shape and are quickly killed if placed in an environment with very high or very low salt concentrations. However, Mycoplasma do have unusually tough membranes that are more resistant to rupture than other bacteria since this cellular membrane has to contend with the host cell factors. Sterols in the membrane contribute to membrane durability by helping to increase the forces that hold the membrane together.
Key Takeaways
- Outside the cell membrane is the periplasm, where enzymes often accumulate that perform degradative and transport functions.
- Outside the periplasm in most microbes is the cell wall. The cell wall is important in giving the cell its shape and preventing the cell membrane from rupturing due to the high osmotic pressure it is under.
- Two general types of cell wall structure are found in bacteria, Gram positive and Gram negative.
- Gram-positive cells have a thick peptidoglycan layer surrounding the cell membrane.
- Gram-negative cells have a thin peptidoglycan layer that is surrounded by a second membrane structure, the outer membrane.
- Finally, there are bacteria that have no cell walls.
Quickcheck 3-5
Warning, you must be logged in to be able to have your exam graded. Answer the questions below and if you are a registered user of the site you will see a Grade Exam button. Click it to have your exam graded.