Latest News
- Clues beginning to emerge on asymtomatic SARS-CoV-2 infection
- Back in November of 2020, during the first wave of the COVID-19 pandemic, I was teaching an in-person microbiology laboratory. One of my students had just been home to see his parents, and they all c…
- Read more
- Could there maybe be better uses of genetics and probiotics?
- Professor Meng Dong and his laboratory have created a probiotic that can metabolize alcohol quickly and maybe prevent some of the adverse effects of alcohol consumption. The scientists cloned a highl…
- Read more
- ChatGPT is not the end of essays in education
- The takeover of AI is upon us! AI can now take all our jobs, is the click-bait premise you hear from the news. While I cannot predict the future, I am dubious that AI will play such a dubious role in…
- Read more
- Fighting infections with infections
- Multi-drug-resistant bacterial infections are becoming more of an issue, with 1.2 million people dying of previously treatable bacterial infections. Scientists are frantically searching for new metho…
- Read more
- A tale of two colleges
- COVID-19 at the University of Wisconsin this fall has been pretty much a non-issue. While we are wearing masks, full in-person teaching is happening on campus. Bars, restaurants, and all other busine…
- Read more
( 42335 Reads)
|Learning Objectives
After reading this section, students will...
- Know that membranes are highly conserved structures found in all living cells and be able to describe their structure.
- Understand the stabilizing forces of membranes.
- Be able to describe the functions of membranes.
- Be able to describe the function and location of the periplasm.
- Be able to describe the function of the cell wall and the two common cell wall structures.
General properties
Membranes define life. The cytoplasmic membrane immediately surrounds the inside of the cell and is perhaps the most conserved structure in living cells. Membranes are thin structures, measuring about 8 nm thick and every living thing on this planet has some type of membrane. They are the major barrier that separates the inside the cell from the outside and allow cells to selectively interact with their environment. Membranes are highly organized and asymmetric. This asymmetry comes from the fact that the membrane that faces the environment performs very different functions than does the side that faces the cytoplasm. Membranes are also dynamic, constantly adapting to changing environmental conditions.
Physical structure
Membranes are composed of lipids and proteins, with about half of all bacteria also containing hopanoids (see section 2-6). Proteins are more numerous in bacterial membranes than in eukaryotic membranes. Bacteria, in general, only contain a single membrane in contact with the cytoplasm, and this has to carry out all the functions of the cell. In eukaryotes, these functions are divided amongst the cytoplasmic membrane and the other organelles.
Some membrane proteins span the membrane (integral or intrinsic membrane proteins) while others sit on the outside or the inside (extrinsic or peripheral membrane proteins). Peripheral membrane proteins easily wash off the membrane using a salt solution, while intrinsic membrane proteins can only be released from it using detergents. Membrane proteins may move within the membrane plane, or the cell may anchor them to structures in or near the membrane.
Much of the general behavior of membranes is dictated by the behavior of lipids in water. Because phospholipids are amphipathic, they congregate when placed in an aqueous environment, similar to how oil will separate from water. This aggregation occurs in a particular way such that the hydrophilic portions face the water and the hydrophobic portions are buried inside. Under the cell's direction, lipids are organized into a bilayer, where there are two sheets of lipids oriented so that the hydrophobic faces of each sheet face each other, as shown in Figure 3.12. Lipid bilayers can be almost any size and can form vesicles spontaneously in water. In the cell, however, the synthesis of membranes is performed by specific enzymes and is tightly controlled.

Figure 3.12. A model of a lipid bilayer. A space-filling representation of a lipid bilayer as developed by H. Heller et al. (J. Phys. Chem. 1993. 97:8343-60).
The cytoplasmic membrane is held together by several forces. Hydrophobic interactions between the alkyl chains of neighboring lipids are a major component of membrane stability. Hydrogen bonds between lipids and between membrane proteins and lipids also hold a membrane together. Further stability comes from negative charges on proteins that form ionic interactions with divalent cations such as Mg+2and Ca+2 and the hydrophilic head of lipids.
The membrane is fluid and has the consistency of a light-grade oil. It has been termed a fluid mosaic: "fluid" because the lipids are free to move about on each side of the membrane and "mosaic" because there is a definite pattern to it. Lipids do not generally switch from outside to inside or vice versa because it is energetically demanding to move the hydrophilic group through the hydrophobic core of the bilayer.
Membranes are a selective barrier
The concentration of solutes, sugars, and most ions is generally much higher within the cell than outside. A fundamental principle of nature is that different concentrations of a given solute tend to equilibrate across a boundary due to diffusion. The hydrophobic region of the membrane provides a critical function: it prevents this diffusion for polar molecules. Therefore, compounds such as amino acids, organic acids, and inorganic salts must be specifically transported across the membrane by proteins, and once inside, these molecules cannot escape. The cell can therefore control the nature and amount of these compounds that enter or leave the cell.
For molecules that are soluble in both the lipid membrane and the surrounding aqueous environment, the law of simple diffusion directs transport. The membrane is not a barrier for such molecules. These types of molecules are uncommon since solubility in both a hydrophobic and a hydrophilic environment is unusual. There is no transport protein for such compounds, so there is no specificity of control or energy cost. The cell cannot create a concentration gradient of these molecules.
There are three basic types of transport systems:
- Facilitated Diffusion
- Group Translocation
- Active Transport
Many of the proteins in the membrane function to help carry out selective transport, particularly of polar compounds. These proteins typically span the entire membrane, making contact with the outside environment and the cytoplasm. They often require the expenditure of energy to help compounds move across the membrane, though cells can also use concentration gradients of these compounds to generate energy, as described below
Facilitated diffusion
This process involves a protein that binds the molecule to be transported and physically moves that compound through the membrane. The molecule's binding to the protein causes a conformational change causing the molecule to face the opposite side from where it was. As shown in Figure 3.14, facilitated diffusion is therefore specific because a protein must bind the molecule. However, these small molecules are readily moved in and out of the cell, so a gradient cannot be formed, nor is energy required.
One example of a class of proteins that carry out facilitated diffusion is aquaporins. Water can diffuse relatively slowly across the membrane, but it was a matter of debate whether this was fast enough for cellular processes. Studies of adaptation to osmotic stress suggested that passive diffusion of water was too slow to explain the rapid changes observed in some bacteria. A search began for the elusive protein that was carrying out this process. Peter Agre discovered the first water channel in 1988. While first described in eukaryotic cells, many living systems are now known to contain them, and the term aquaporins was coined to describe them. Over 100 aquaporins are present in bacterial systems, and while these do not share a high degree of sequence homology with eukaryotic aquaporins, they share a significant structural similarity. There are two large classes of aquaporins: those that only allow water to flow through them and those that will also transport glycerol and a few other small uncharged molecules. Aquaporins are not present in every microorganism; some species appear to lack them. In these cases, the microbe may rely solely on diffusion across the membrane, or its aquaporin has a novel sequence.
Aquaporins (Figure 3.13) are intrinsic membrane proteins of about 23 kDa that contain six α-helices arranged around a central core channel. Each aquaporin will associate with three others to form a homotetramer in the membrane. The central channel is narrow and contains a conserved structure that can selectively allow only the passage of the desired molecule. In this way, water can rapidly flow into and out of the cell, speeding up its transport through the membrane.
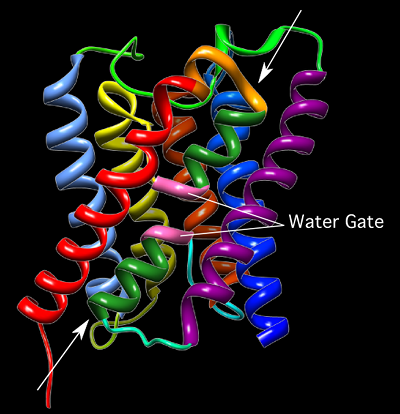
Figure 3.13. Aquaporin. A ribbon diagram of the 23 kDA Aquaporin from E. coli. Each of the 6 α helices that line the water channel are shown. The arrows indicate the path of water through the protein. The two sequences of 3 amino acids (Asparagine-Proline-Alanine) are shown in pink. This area is thought to be important in discriminating, only allowing water molecules to pass through the pore. Thus, serving as a gate.
While aquaporins are essential to cell function, they also create a serious problem. The inside of the cell is full of many types of solutes: proteins, nucleic acids, other small molecules, and ions. In comparison, the outside environment, in most cases, is very dilute. Because of this, there is a higher concentration of water outside the cell than inside the cell. Nature hates imbalances such as this, and to correct the issue, water tends to flow into the cell by a process called osmosis. Osmosis causes high pressure against the cell membrane. This pressure would rapidly cause lysis of most cells, and one of the major purposes of the peptidoglycan of the cell wall (discussed in a coming section) is to prevent the cell membrane from bursting. Figure 3.14 shows an animation of facilitated diffusion.
Figure 3.14. Facilitated Diffusion. An animation of the migration of solutes in and out of the cell as facilitated by a protein. Notice that this mechanism does not lead to a solute concentration inside the cell that is higher than outside. Rather, it leads to an equilibrium of that solute across the gradient.
Group translocation
In this process, a protein specifically binds the target molecule and transports it inside the cell while simultaneously modifying it chemically. Most group translocations require energy and tend to be unidirectional, unlike facilitated diffusion. The substrates of catabolic pathways, such as sugars, are sometimes transported by group translocation. Group translocation is an efficient way to both bring substrate into the cell and begin the breakdown process. Figure 3.15 shows an animation of group translocation.
Figure 3.15. Group Translocation. An animation of group translocation. The glucose molecule that is being transported into the cell is modified by the addition of a phosphate from phosphoenolpyruvate to form glucose-6-phosphate.
Active transport
In active transport, energy is expended to transport small molecules without chemically altering the substrate. The process is efficient enough to cause the internal concentration in the cell to reach many times its external concentration. Active transport proteins are molecular pumps that expend energy to pump their substrates against a concentration gradient. This energy comes in two forms: ATP and ion gradients. In ATP-based active transport, the transport proteins couples ATP hydrolysis to the movement of the small molecule across the membrane. The animation in Figure 3.16 shows an animation of an ABC transporter. One large group of proteins involved in this type of transport is the ATP binding cassette (ABC) transporters. ABC transporters are present in all living species, with 80 identified in the E. coli genome and 48 in the human genome. An example of an ABC transporter is the maltose-binding protein of E. coli.
Figure 3.16. The Maltose Transporter, an ATP Binding Cassette Transporter. Three proteins are involved in the maltose transporter. A maltose-binding protein in the periplasm and two membrane-bound transporter proteins. One serves at the gate, while the other binds and hydrolyzes ATP. The hydrolysis of ATP to ADP changes the conformation of the gate, allowing the passage of maltose into the cell.
Ion gradient active transport uses the energy of one chemical gradient, that of the specific ion, to drive the creation of a different gradient, the uptake of the small molecule. The ion gradient that supports the work is at a higher concentration on one side of the membrane than the other. The transport protein both binds its small molecule to transport and provides a gateway for this ion to fall down its concentration gradient. When the ion moves through its gateway, it causes a conformational change in the protein, and this change helps to transport the target small molecule into the cell.
Active transport proteins may be highly specific for only one molecule or may be able to carry a class of chemically related molecules. An example of a more general transport protein is the branch chain amino acid transporter of Pseudomonas aeruginosa, which transports leucine, valine, and isoleucine. Table 3.1 summarizes the various properties of transport mechanisms.
Table 3.1. Properties of various transport systems
| Property | Passive Diffusion | Facilitated Diffusion | Active Transport | Group Translocation |
| Carrier Mediated | - | + | + | + |
| Concentration Against Gradient | - | - | + | Not Applicable |
| Specificity | - | + | + | + |
| Energy Expended | - | - | + | + |
| Solute Modified During Transport | - | - | - | + |
A comparison of the methods for transporting molecules through the membrane and into the cell.
Membranes can help generate energy
Many cells use respiratory processes to obtain their energy. During respiration, organic or inorganic compounds that contain energy are oxidized, releasing electrons to do work. In many microorganisms, these electrons find their way to the membrane, passing through a series of electron carriers, as shown in Figure 3.17. During this operation, protons move outside the cell. This movement creates a gradient of protons across the cell membrane, energizing it in a fashion similar to charging a battery. The energy of this gradient can then be used to do work directly, a process known as the proton motive force, or can be channeled into a special protein known as ATP synthase. ATP synthase can convert: ADP to ATP, and the ATP can itself do work.
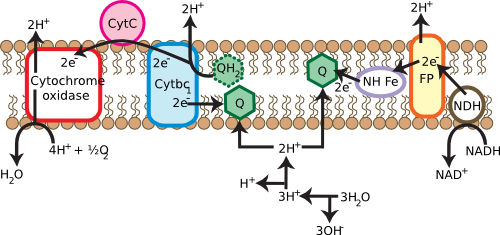
Figure 3.17. Generating the proton motive force. Membranes are critical in many cells for the generation of usable energy. The cartoon shows the various membrane proteins involved in converting high-energy electrons from photoreceptors into useful energy. They do so by forming a proton gradient across the cell membrane, termed a proton motive force, which is in turn used by other proteins to synthesize ATP. This will be discussed in greater detail in the chapter on metabolism.
The prokaryotic cells performing photosynthesis also have a membrane system that does a very similar process. Light excites electrons found in pigmented proteins in the membrane, and the electrons again move through a series of electron carriers. As above, a proton motive force is generated and used to synthesize ATP. The chapters on metabolism and photosynthesis discuss the specifics of these systems.
Membranes are also important in their own synthesis and can fold inward for specialized functions
Membranes also contain specialized enzymes that carry out certain biosynthetic functions. For example, the last few steps of lipid synthesis take place inside the membrane. Another example is cell wall synthesis and assembly. Much of the synthesis of cell wall monomers occurs there, and the stitching together of the cell wall polymer takes place while it is anchored to the membrane. In addition, any cellular protein that carries out its function outside the cell membrane (such as outer membrane and extracellular proteins) must pass through that membrane. During their synthesis, the cytoskeleton guides the ribosome to the cytoplasmic face of the membrane, and the growing peptide chain is synthesized directly into the lipid bilayer. Integral membrane proteins then fold up and stay in the membrane while extracellular proteins move through the membrane and take on their final shape on the other side.
Infoldings of the membrane are found in some photosynthetic bacteria. These bacteria use pigments in their membranes to capture light energy. Under low light, they need to increase the surface area to catch more light. They cannot make the membrane thicker, but they can increase the surface area by creating regions where the membrane folds into the cytoplasm. These invaginations are still attached to the cytoplasmic membrane and a picture of such structures, termed the intracytoplasmic membrane in the case of Rhodobacter sphaeroides, is shown in Figure 3.18.
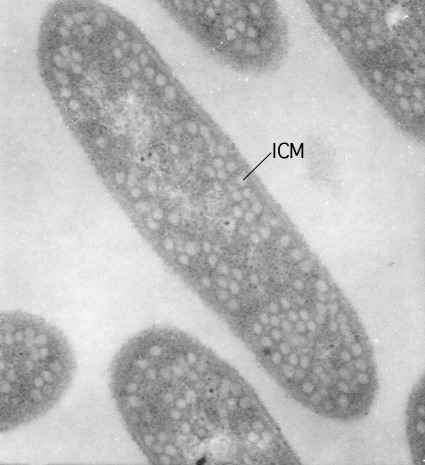
Figure 3.18. The intracytoplasmic membrane of Rhodobacter sphaeroides.. This electron micrograph shows the complicated infolding of the cytoplasmic membrane of R. sphaeroides when it is performing photosynthesis. This infolding creates a larger membrane surface area into which light-harvesting complexes can be inserted. Under low-light conditions many light-harvesting complexes are needed to capture the small number of photons striking the microbe. (Source: Samuel Kaplan, University of Texas - Houston Medical School)
Key Takeaways
- Membranes are some of the most conserved structures in biology. They are composed of mostly lipids, with some proteins and polysaccharides. Their characteristics are dictated by the amphipathic nature of lipids, containing a hydrophilic exterior and a hydrophobic core.
- Membranes serve as a selective barrier, preventing the passing of all but very non-polar molecules.
- To facilitate transport, membranes will contain transport proteins that move selected molecules across.
- The methods used by transport proteins can be classified as facilitated diffusion, group translocation and active transport.
- Group translocation and active transport concentrate molecules against a gradient and require energy.
- Membrane are also involved in the generation of energy and participate in their own synthesis.
Quickcheck 3-3
Warning, you must be logged in to be able to have your exam graded. Answer the questions below and if you are a registered user of the site you will see a Grade Exam button. Click it to have your exam graded.